
СП 124.13330.2012 «СНиП 41-02-2003 Тепловые сети» — DWGFORMAT
Область применения
Свод правил распространяется на тепловые сети (со всеми сопутствующими конструкциями) от выходных запорных задвижек (исключая их) коллекторов источника теплоты или от наружных стен источника теплоты до выходных запорных задвижек (включая их) центральных тепловых пунктов и до входных запорных органов индивидуальных тепловых пунктов (узлов вводов) зданий (секции зданий) и сооружений, транспортирующие горячую воду с температурой до 200 градусов Цельсия и давлением до 2,5 МПа включительно, водяной пар с температурой до 440 градусов Цельсия и давлением до 6,3 МПа включительно, конденсат водяного пара.
Содержание
1 Область применения
2 Нормативные ссылки
3 Термины и определения
4 Классификация
5 Общие положения
6 Схемы теплоснабжения и тепловых сетей
7 Теплоносители и их параметры
8 Гидравлические режимы
9 Трассы и способы прокладки тепловых сетей
10 Конструкция трубопроводов
11 Тепловая изоляция
13 Защита трубопроводов от коррозии
14 Тепловые пункты
15 Электроснабжение и система управления
16 Дополнительные требования к проектированию тепловых сетей в особых природных и климатических условиях строительства
17 Энергоэффективность тепловых сетей
Приложение А (обязательное).
 Расстояния от строительных конструкций тепловых сетей или оболочки изоляции трубопроводов при бесканальной прокладке до зданий, сооружений и инженерных сетей
Расстояния от строительных конструкций тепловых сетей или оболочки изоляции трубопроводов при бесканальной прокладке до зданий, сооружений и инженерных сетейПриложение Б (обязательное). Требования к размещению трубопроводов при их прокладке в непроходных каналах, тоннелях, надземной и в тепловых пунктах
Приложение В (рекомендуемое). Удельные показатели максимальной тепловой нагрузки на отопление и вентиляцию жилых домов
Приложение Г (рекомендуемое). Нормы расхода горячей воды потребителями и удельная часовая величина теплоты на ее нагрев
Приложение Д (обязательное). Мероприятия, обеспечивающие безопасность эксплуатации
Приложении Е (обязательное). Требования к качеству сетевой и подпиточной воды тепловых сетей
Библиография
Поделиться в социальных сетях
Ещё записи из рубрики «» Новый мост в городе Чэнду В китайском городе Чэнду возвели необычный пешеходный мост. Сооружение называется «Учацзы» («Пять. ..
..
Проектирование тепловых сетей
Тепловая сеть – совокупность устройств (центральные тепловые пункты, насосные станции) и система соединенных между собой участков теплопроводов, по которым с помощью теплоносителя (пар или горячая вода) тепловая энергия передается (транспортируется) от источников тепловой энергии до теплопотребляющих установок и далее к конечным потребителям.
Общая протяженность тепловых сетей в России в двухтрубном исчислении составляет около 183 300 км (данные 2000 года).
Россия один из лидеров стран производителей тепловой энергии. Около 45% мирового производства тепловой энергии приходится на Россию. Система теплоснабжения России состоит из 50000 локальных систем теплоснабжения, которые обслуживает 17000 предприятий. За выработку тепловой энергии отвечает 526 ТЭЦ и 72000 котельных. На сегодняшний день в России централизованным теплоснабжением обеспечено более 80% жилищного фонда, а 64% населения обеспечено горячей водой. В отрасли теплоснабжения и связанных с ним структур работает более 2 миллионов человек.
В отрасли теплоснабжения и связанных с ним структур работает более 2 миллионов человек.
Мы предлагаем комплексное проектирование тепловых сетей, включающее в себя технико-экономическое обследование существующих систем теплоснабжения и оптимизация схем теплоснабжения с учетом всех энергосберегающих факторов.
Наша организация имеет богатый опыт работы в сфере проектирования тепловых сетей, мы осуществляли такие виды работ как:
- Вынос тепловых сетей из пятна застройки строящегося здания.
- Подключение к теплоснабжению как индивидуальных потребителей, так и крупных застроек.
- Реконструкция тепловых сетей.
- Осуществление временного теплоснабжения.
При выполнении работ особое внимание уделяется оптимизации затрат на строительство проектируемой тепловой сети, а также ее надежности и качеству.
В проектах применяются проверенные в эксплуатации стальные трубы в ППУ изоляции, а также трубы из сшитого полиэтилена “Изопрофлекс-А”. Тепловые сети проходят обязательную проверку на прочность в программе “СТАРТ”,широко применяется бесканальная прокладка и естественная компенсация. Все технические решения по проектам согласовываются в эксплуатирующих и надзорных организациях.
Тепловые сети проходят обязательную проверку на прочность в программе “СТАРТ”,широко применяется бесканальная прокладка и естественная компенсация. Все технические решения по проектам согласовываются в эксплуатирующих и надзорных организациях.
Мы предлагаем комплексное проектирование тепловых сетей, включающее в себя технико-экономическое обследование существующих систем теплоснабжения и оптимизация схем теплоснабжения с учетом всех энергосберегающих факторов.
- Проектирование городских магистральных тепловых сетей, разводящих и внутриквартальных сетей.
- Проектирование тепловых камер.
- Проектирование и разработка индивидуальных тепловых узлов.
- Проектирование тепловых сетей различных типов прокладки (закрытый, открытый и т.д.)
Вся проектная документация тепловых сетей разрабатывается специалистами компании АТ-СТРОЙГРУПП
в строгом соответствии следующими нормативам:
- СНиП 207-01-89* «Градостроительство.
 Планировка и застройка городов, посёлков и сельских населенных пунктов. Нормы проектирования».
Планировка и застройка городов, посёлков и сельских населенных пунктов. Нормы проектирования». - СНиП 41-02-2003 «Тепловые сети».
- СНиП 41-02-2003 «Тепловая изоляция оборудования и трубопроводов».
- СНиП 3.05.03-85 «Тепловые сети» (предприятие тепловых сетей).
- ГОСТ 21-605-82 «Тепловые сети (тепломеханическая часть)».
- СНиП 2.04.03-85 «Канализация. Наружные сети и сооружения».
- СНиП 3.05.04-85 «Наружные сети и сооружения водоснабжения и канализации».
- Правила подготовки и производства земляных работ, устройства и содержания строительных площадок в городе Москве, утверждённых постановлением правительства г .Москвы №857-ПП от 07.12.2004г.
- ПБ 10-573-03 «Правила устройства и безопасной эксплуатации трубопроводов пара и горячей воды».
СНиП 41-02-2003 Тепловые сети
ТЕПЛОВАЯ ИЗОЛЯЦИЯ
11.1 Для тепловых сетей следует, как правило, принимать теплоизоляционные материалы и конструкции, проверенные практикой эксплуатации. Новые материалы и конструкции допускаются к применению при положительных результатах независимых испытаний, проведенных специализированными лабораториями.
Новые материалы и конструкции допускаются к применению при положительных результатах независимых испытаний, проведенных специализированными лабораториями.
11.2 Материалы тепловой изоляции и покровного слоя теплопроводов должны отвечать требованиям СНиП 41-03, норм пожарной безопасности и выбиратъся в зависимости от конкретных условий и способов прокладки.
При совместной подземной прокладке в тоннелях (проходных каналах) теплопроводов с электрическими или слаботочными кабелями, трубопроводами, транспортирующими горючие вещества, не допускается применять тепловую изоляционную конструкцию из горючих материалов. При отдельной прокладке теплопроводов в тоннелях (проходных каналах) применение негорючих материалов (НГ) обязательно только для покровного слоя тепловой изоляции теплопроводов.
При надземной прокладке теплопроводов рекомендуется применять для покровного слоя теплоизоляции негорючие материалы и групп горючести Г1 и Г2.
При подземной бесканальной прокладке и в непроходных каналах допускается применять горючие материалы теплоизоляционного и покровного слоев.
11.3 Тоннель (проходной канал) следует разделять через каждые 200 м на отсеки противопожарными перегородками 1-го типа с противопожарными дверями 2-го типа.
11.4 При прокладке теплопроводов в теплоизоляции из горючих материалов следует предусматривать вставки из негорючих материалов длиной не менее 3 м:
в каждой камере тепловой сети и на вводе в здания;
при надземной прокладке — через каждые 100 м, при этом для вертикальных участков через каждые 10 м;
в местах выхода теплопроводов из грунта.
При применении конструкций теплопроводов в теплоизоляции из горючих материалов в негорючей оболочке допускается вставки не делать.
11.5 Детали крепления теплопроводов должны выполняться из коррозионно- стойких материалов или покрываться антикоррозионными покрытиями.
11.6 Выбор материала тепловой изоляции и конструкции теплопровода следует производить по экономическому оптимуму суммарных эксплуатационных затрат и капиталовложений в тепловые сети, сопутствующие конструкции и сооружения. При выборе теплоизоляционных материалов, применение которых вызывает необходимость изменения параметров теплоносителя (расчетной температуры, режимов регулирования и т.п.), следует производить сопоставление вариантов систем централизованного теплоснабжения в целом.
При выборе теплоизоляционных материалов, применение которых вызывает необходимость изменения параметров теплоносителя (расчетной температуры, режимов регулирования и т.п.), следует производить сопоставление вариантов систем централизованного теплоснабжения в целом.
Выбор толщины теплоизоляции следует производить по СНиП 41-03 на заданные параметры с учетом климатологических данных пункта строительства, стоимости теплоизоляционной конструкции и теплоты.
11.7 При определении тепловых потерь трубопроводами расчетная температура теплоносителя принимается для подающих теплопроводов водяных тепловых сетей:
при постоянной температуре сетевой воды и количественном регулировании — максимальная температура теплоносителя;
при переменной температуре сетевой воды и качественном регулировании среднегодовая температура теплоносителя 110 °С при температурном графике регулирования 180 — 70 °С, 90 °С при 150 — 70 °С, 65 °С при 130 — 70 °С и 55 °С при 95 -70 °С. Среднегодовая температура для обратных теплопроводов водяных тепловых сетей принимается 50 °С.
11.8 При размещении теплопроводов в служебных помещениях, технических подпольях и подвалах жилых зданий температура внутреннего воздуха принимается равной 20 °С, а температура на поверхности конструкции теплопроводов не выше 45 °С.
11.9 При выборе конструкций теплопроводов надземной и канальной прокладки следует соблюдать требования к теплопроводам в сборке:
при применении конструкций с негерметичными покрытиями покровный слой теплоизоляции должен быть водонепроницаемым и не препятствовать высыханию увлажненной теплоизоляции.
при применении конструкций с герметичными покрытиями обязательно устройство системы оперативного дистанционного контроля (ОДК) увлажнения теплоизоляции;
показатели температуростойкости, противостояния инсоляции должны находиться в заданных пределах в течение всего расчетного срока службы для каждого элемента или конструкции;
скорость наружной коррозии стальных труб не должна превышать 0,03 мм/год.
11.10 При выборе конструкций для подземных бесканальных прокладок тепловых сетей следует рассматривать две группы конструкций теплопроводов:
группа «а» — теплопроводы в герметичной паронепроницаемой гидрозащитной оболочке. Представительная конструкция — теплопроводы заводского изготовления в пенополиуретановой теплоизоляции с полиэтиленовой оболочкой по ГОСТ 30732;
Представительная конструкция — теплопроводы заводского изготовления в пенополиуретановой теплоизоляции с полиэтиленовой оболочкой по ГОСТ 30732;
группа «б» — теплопроводы с паропроницаемым гидрозащитным покрытием или в монолитной теплоизоляции, наружный уплотненный слой которой должен быть водонёпроницаемым и одновременно паропроницаемым, а внутренний слой, прилегающий к трубе, — защищать стальную трубу от коррозии. Представительные конструкции — теплопроводы заводского изготовления в пенополимерминеральной или армопенобетонной теплоизоляции.
11.11 Обязательные требования к теплопроводам группы «а»:
равномерная плотность заполнения конструкции теплоизоляционным материалом;
герметичность оболочки и наличие сйстемы ОДК, организация замены влажного участка сухим;
показатели температуроcтойкости должны находиться в заданных пределах в течение расчетного срока службы;
скорость наружной коррозии труб не должна превышать 0,03 мм/год;
стойкость к истиранию защитного покрытия . — на более 2 мм/25 лет.
Обязательные требования к физико-техническим характеристикам конструкций теплопроводов группы «б»:
показатели температуростойкости должны находиться в заданных пределах в течение расчетного срока службы; скорость наружной коррозии стальных труб не должна превышать 0,03 мм/год.
11.12 При расчете толщины изоляции и определении годовых потерь теплоты теплопроводами, проложенными бесканально на глубине заложения оси теплопровода более 0,7 м, за расчетную температуру окружающей среды принимается средняя за год температура грунта на этой глубине.
При глубине заложения теплопровода от верха теплоизоляционной конструкции менее 0,7 м за расчетную температуру окружающей среды принимается та же температура наружного воздуха, что и при надземной прокладке.
Для определения температуры грунта в температурном поле подземного теплопровода температура теплоносителя должна приниматься:
для водяных тепловых сетей — по температурному графику регулирования при средней месячной температуре наружного воздуха расчетного месяца;
для сетей горячего водоснабжения — по максимальной температуре горячей воды.
11.13 При выборе конструкций надземных теплопроводов следует учитывать следующие требования к физико-техническим характеристикам конструкций теплопроводов:
показатели температуростойкости должны находиться в заданных пределах в течение расчетного срока службы конструкции; скорость наружной коррозии стальных труб не должна превышать 0,03 мм/год.
11.14 При определении толщины теплоизоляции теплопроводов, проложенных в проходных каналах и тоннелях, следует принимать температуру воздуха в них не более 40 °С.
11.15 При определении годовых потерь теплоты теплопроводами, проложенными в каналах и тоннелях, параметры теплоносителя следует принимать по 11.7.
11.16 При прокладке тепловых сетей в не-проходных каналах и бесканально коэффициент теплопроводности теплоизоляции должен приниматься с учетом возможного увлажнения конструкции теплопроводов.
СКАЧАТЬ СНиП 41-02-2003 Тепловые сети
СП 124.13330.2012 | СНиП Тепловые сети
Здравствуйте, друзья! Сегодня решил пополнить рубрику «Библиотека теплоэнергетика» очередной статьей. Речь пойдет о СП 124.13330.2012, ТЕПЛОВЫЕ СЕТИ, актуализированная редакция СНиП 41-02-2003. Данный свод правил был введен в действие с 01 января 2013 года. Итак, о чем же этот свод правил, и чем он может быть полезен?
Речь пойдет о СП 124.13330.2012, ТЕПЛОВЫЕ СЕТИ, актуализированная редакция СНиП 41-02-2003. Данный свод правил был введен в действие с 01 января 2013 года. Итак, о чем же этот свод правил, и чем он может быть полезен?
В пункте 1 «Область применения» СП можно прочитать, что этот нормативный документ применяется при проектировании новых и реконструкции, модернизации тепловых сетей централизованного теплоснабжения. В п.2 «Нормативные ссылки» приведены ссылки на нормативные документы, которые упоминаются в своде правил. Следующий пункт, третий – это «Термины и определения». Я в теплоэнергетике работаю уже более 15 лет, и многие термины понимаю по своему, здесь же можно прочитать, как эти термины будут звучать правильно.
СНиП 41-02-2003 Тепловые сети можно скачать здесь:
СНиП 41-02-2003, Тепловые сети
СП 124.13330.2012, ТЕПЛОВЫЕ СЕТИ можно скачать здесь:
СП 124.13330.2012 Тепловые сети
В четвертом пункте «Классификация» — самое интересное это то, как потребители делятся на категории по степени надежности теплоснабжения. Всего категорий три. Потребители, для которых не допускается снижение температуры воздуха в помещениях; потребители, для которых допускается временное снижение внутренней температуры; и третья категория – иные потребители.
Всего категорий три. Потребители, для которых не допускается снижение температуры воздуха в помещениях; потребители, для которых допускается временное снижение внутренней температуры; и третья категория – иные потребители.
Пункт № 5 «Общие положения» сообщает о том, какие в своде правил установлены требования по различным параметрам – безопасности, надежности, обеспечению, а также другие общие определения. Пункт шестой – это «Схемы теплоснабжения и тепловых сетей». Здесь очень много полезной информации о магистральных тепловых сетях, подпитке, надежности, резервировании, живучести, сборе и возврате конденсата. Далее в своде правил — п.7 «Теплоносители и их параметры». Очень нужный и полезный пункт, здесь в частности, говорится о том, какие виды регулирования следует применять на теплоисточнике, ЦТП, ИТП, о том какие температуры применять для расчета температурного графика.
Пункт № 8 «Гидравлические режимы». Здесь говорится о том, какие виды гидравлических режимов предусматриваются, какие условия следует принимать при расчете режимов.
Очень важный и нужный пункт № 11 «Тепловая изоляция». Знать нормативные требования к тепловой изоляции теплосетей необходимо. Следующим пунктом идет двенадцатый – «Строительные конструкции». Здесь читаем о том, что такое подземная и надземная прокладка тепловых сетей, нормы к ним. П. № 13 «Защита трубопроводов от коррозии». Надо сказать, что для стальных трубопроводов – коррозия главный враг. Причем коррозия происходит как снаружи, так и изнутри. В этом пункте говорится о том, какие виды защиты следует применять для трубопроводов тепловых сетей. Следующий по списку – номер четырнадцать, «Тепловые пункты». Здесь читаем о том, какие нормы и правила применяются к тепловым пунктам тепловых сетей.
Следующий по списку – номер четырнадцать, «Тепловые пункты». Здесь читаем о том, какие нормы и правила применяются к тепловым пунктам тепловых сетей.
П. № 15 – «Электроснабжение и система управления». В числе прочего, в этом пункте говорится о диспетчеризации управления на тепловых сетях. Важность этого мероприятия трудно переоценить. Название пункта № 16 «Дополнительные требования к проектированию тепловых сетей в особых природных и климатических условиях строительства» говорит само за себя. Тем более что страна у нас большая, и таких географических районов немало. И наконец, последний пункт свода правил – это пункт № 17 «Энергоэффективность тепловых сетей». Пункт немаловажный в современных условиях рыночной экономики.
В качестве приложений к своду правил 124.13330.2012, ТЕПЛОВЫЕ СЕТИ – приложение А «РАССТОЯНИЯ ОТ СТРОИТЕЛЬНЫХ КОНСТРУКЦИЙ ТЕПЛОВЫХ СЕТЕЙ ИЛИ ОБОЛОЧКИ ИЗОЛЯЦИИ ТРУБОПРОВОДОВ ПРИ БЕСКАНАЛЬНОЙ ПРОКЛАДКЕ ДО ЗДАНИЙ, СООРУЖЕНИЙ И ИНЖЕНЕРНЫХ СЕТЕЙ»,
приложение Б «ТРЕБОВАНИЯ К РАЗМЕЩЕНИЮ ТРУБОПРОВОДОВ ПРИ ИХ ПРОКЛАДКЕ В НЕПРОХОДНЫХ КАНАЛАХ, ТОННЕЛЯХ, НАДЗЕМНОЙ И В ТЕПЛОВЫХ ПУНКТАХ», приложение В «УДЕЛЬНЫЕ ПОКАЗАТЕЛИ МАКСИМАЛЬНОЙ ТЕПЛОВОЙ НАГРУЗКИ НА ОТОПЛЕНИЕ И ВЕНТИЛЯЦИЮ ЖИЛЫХ ДОМОВ, Вт/м2»,
приложение Г «НОРМЫ РАСХОДА ГОРЯЧЕЙ ВОДЫ ПОТРЕБИТЕЛЯМИ И УДЕЛЬНАЯ ЧАСОВАЯ ВЕЛИЧИНА ТЕПЛОТЫ НА ЕЕ НАГРЕВ»,
приложение Д «МЕРОПРИЯТИЯ, ОБЕСПЕЧИВАЮЩИЕ БЕЗОПАСНОСТЬ ЭКСПЛУАТАЦИИ»,
приложение Е «ТРЕБОВАНИЯ К КАЧЕСТВУ СЕТЕВОЙ И ПОДПИТОЧНОЙ ВОДЫ ТЕПЛОВЫХ СЕТЕЙ».
На этом обзор СП 124.13330.2012, ТЕПЛОВЫЕ СЕТИ будем считать законченным.
Нормативные документы в строительстве и эксплуатации сетей отопления, теплоснабжения, вентиляции и кондиционирования
В Украине при строительстве и эксплуатации сетей отопления, теплоснабжения, вентиляции и кондиционирования действуют следующие нормативные документы:
1. ДСТУ Б В.2.7-144:2007 «Труби для мереж холодного та гарячого водопостачання із поліпропілену. Технічні умови»
2. Изготовление соединительных деталей для водопроводных полипропиленовых труб производится согласно требованиям ДСТУ Б В.2.5-18-2001. Детали соединительные из полипропилена для сварки нагретым инструментом враструб при строительстве систем холодного и горячего водоснабжения. Технические условия — скачать
3. Изготовление водопроводных труб из сшитого полиэтилена производится согласно требованиям ДСТУ Б В.2.5-17-2001.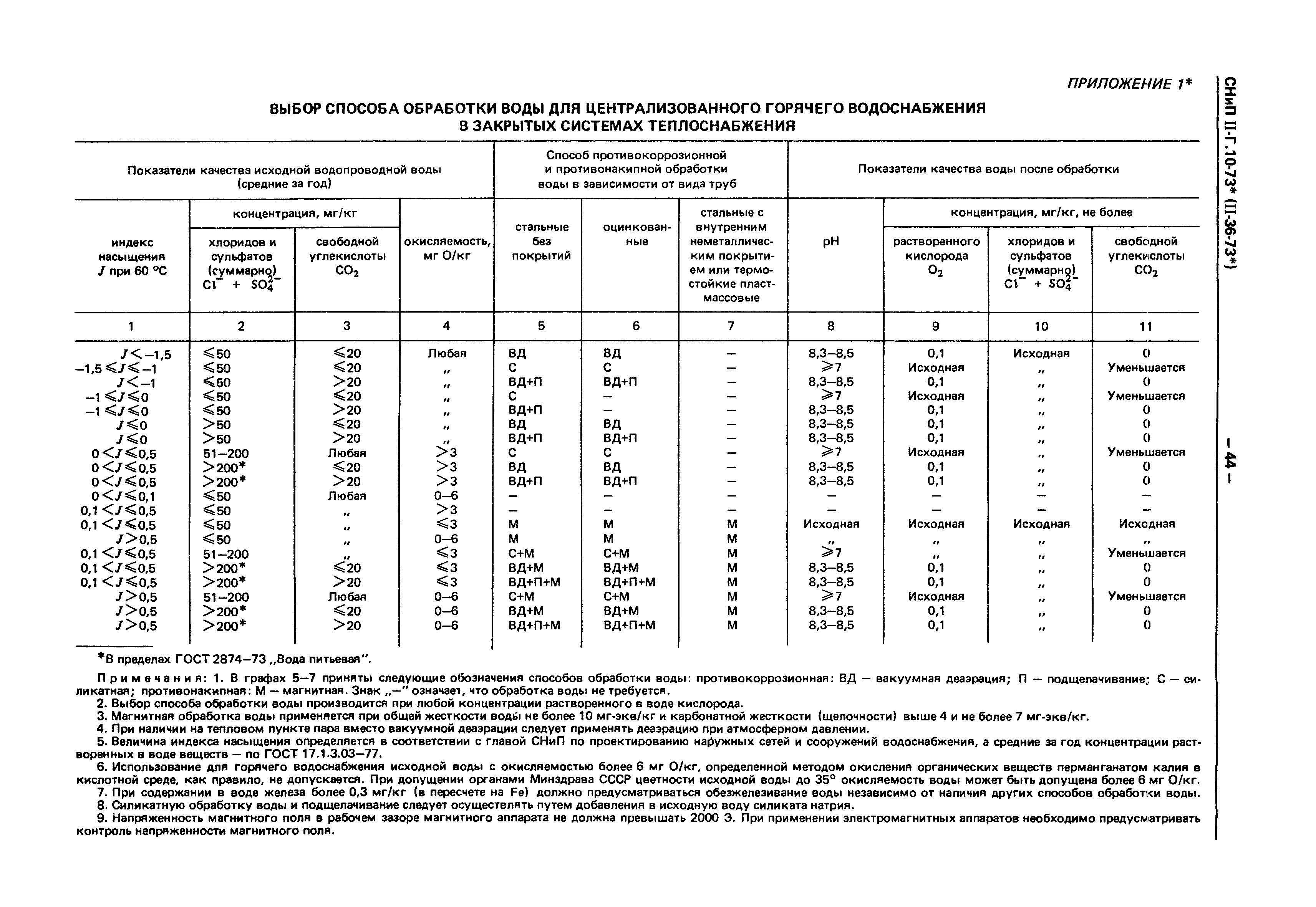 Трубы из структурированного полиэтилена для систем холодного, горячего водоснабжения и отопления. Технические условия — скачать
Трубы из структурированного полиэтилена для систем холодного, горячего водоснабжения и отопления. Технические условия — скачать
4. Изготовление водопроводных труб из сшитого полиэтилена с тепловой изоляцией производится согласно требованиям ДСТУ Б В.2.5-21-2002. Трубы из структурированного полиэтилена с тепловой изоляцией из вспененного полиэтилена и защитной гофрированной полиэтиленовой оболочкой для сетей холодного, горячего водоснабжения и водяного отопления. Технические условия — скачать
5. СНиП 2.04.05-91. Строительные нормы и правила. Отопление, вентиляция и кондиционирование — скачать
6. СНиП 2.04.07-86. Строительные нормы и правила. Тепловые сети (в редакции 1988 года) — скачать
7. СНиП 2.04.14-88. Строительные нормы и правила. Тепловая изоляция оборудования и трубопроводов — скачать
8. СНиП 3.05.03-85. Строительные нормы и правила. Тепловые сети — скачать
9. СНиП II-3-79. Строительные нормы и правила. Строительная теплотехника — скачать
Строительные нормы и правила. Строительная теплотехника — скачать
10. СНиП II-58-75. Строительные нормы и правила. Электростанции тепловые — скачать
В Российской Федерации действуют следующие нормативные документы:
1. СНиП 41-01-2003. Строительные нормы и правила. Отопление, вентиляция и кондиционирование — скачать
2. СНиП 41-02-2003. Строительные нормы и правила. Тепловые сети — скачать
3. СНиП 41-03-2003. Строительные нормы и правила. Тепловая изоляция оборудования и трубопроводов — скачать
4. СП 41-101-95. Свод правил. Проектирование тепловых пунктов — скачать
5. СП 41-102-2000. Свод правил по проектированию и строительству. Проектирование и монтаж трубопроводов систем отопления с использованием металлополимерных труб — скачать
6. СП 41-103-2000. Свод правил. Проектирование тепловой изоляции оборудования и трубопроводов — скачать
7. СП 41-104-2000. Свод правил. Проектирование автономных источников теплоснабжения — скачать
СП 41-104-2000. Свод правил. Проектирование автономных источников теплоснабжения — скачать
8. СП 41-108-2004. Свод правил. Поквартирное теплоснабжение жилых зданий с теплогенераторами на газовом топливе — скачать
9. СП 13-101-99. Свод правил. Правила надзора, обследования, проведения технического обслуживания и ремонта промышленных дымовых и вентиляционных труб — скачать
10. СП 23-02-2003. Свод правил. Тепловая защита зданий — скачать
11. СП 23-101-2000. Свод правил по проектированию и строительству. Проектирование тепловой защиты зданий — скачать
12. РЕКОМЕНДАЦИИ по проектированию энергоэкономичных технических решений систем отопления, вентиляции и водоснабжения встроенно-пристроенных в жилые здания помещений общественного назначения — скачать
теплотрассы, теплосети по СНиП, сколько метров в каждую сторону при подземной прокладке
Охранная зона тепловых сетей – территория, согласно нормам СНиП (СП), необходимая для предотвращения повреждений трубопровода при проведении посторонних работ, осуществления строительства, высадки зеленых насаждений. Она используется при прокладке коммуникаций других сетей (электрических и газовых). Охранная зона теплосети – это пространство со своим собственным правовым режимом, запрещающим или ограничивающим любые виды хозяйственной деятельности, не имеющие отношения к теплоснабжению потребителя. Это не только территория с особым режимом эксплуатации, на которой действуют нормативные документы и специальные постановления.
Она используется при прокладке коммуникаций других сетей (электрических и газовых). Охранная зона теплосети – это пространство со своим собственным правовым режимом, запрещающим или ограничивающим любые виды хозяйственной деятельности, не имеющие отношения к теплоснабжению потребителя. Это не только территория с особым режимом эксплуатации, на которой действуют нормативные документы и специальные постановления.
Надземная тепловая сеть
Основные нормативные документы
В октябре 2019 года планируется проведение отраслевой конференции по теплоснабжению, на которой в числе актуальных проблем будет рассматриваться и правовая сторона вопроса. Владельцам тепловых сетей постоянно приходится сталкиваться с трудностями эксплуатации и невозможностью полноценного ремонта.
Ремонт тепловых магистралей в канале
Это происходит из-за того, что постоянно нарушается охранная зона теплотрассы. Над подземными сетями устраиваются стоянки автомобилей, высаживаются зеленые насаждения – кусты и деревья, возводятся временные сооружения вроде ларьков и гаражей.
Правовое понятие «зона тепловых сетей» имеет под собой весомое юридическое обеспечение.
Несоблюдение режима охранных зон карается финансовыми и административными санкциями. В недалеком будущем планируется усиление мер по обеспечению прав собственников тепловых сетей.
Задвижки трубопровода
Охранная зона, или санитарно-защитная зона, создается не только для теплосетей. Также она нужна для любого трубопровода, транспортных магистралей (в том числе и железнодорожных), объектов транспортировки электроэнергии.
У каждой отрасли имеются свои документы и правила, постановления правительства и профильных министерств, приказы и особые распоряжения. Все это необходимо для корректной эксплуатации, предотвращения повреждений сооружений. А также во избежание потенциального вреда здоровью граждан, оказавшихся в зоне аварии.
Наружная теплосеть
Охранная зона сетей имеет свой нормативный пакет документов, из которых основополагающими считаются:
- Приказ Госстроя Российской Федерации за № 92 «Организационно-методические рекомендации по эксплуатации теплосетей» и собственно рекомендации, которые направлены на оптимизацию и безопасность коммуникаций по теплопоставкам.

- Типовые правила эксплуатации, утвержденные в августе 1992 года Приказом Министерства архитектуры и строительства за № Они обязательны к выполнению любыми организациями, в компетенции которых входит строительная, реконструкционная и ремонтная деятельность в области теплотрассы и на участке, определенном в правилах как охранная зона теплотрассы.
- В субъектах федерации, регионах и населенных пунктах на основании данных типовых правил и утвержденных правительством СНиП (СП) могут разрабатываться и утверждаться местные инструкции. Их требования не могут быть меньше, чем указанные в Типовых правилах и соответствующем СНиП.
Подземная тепловая магистраль
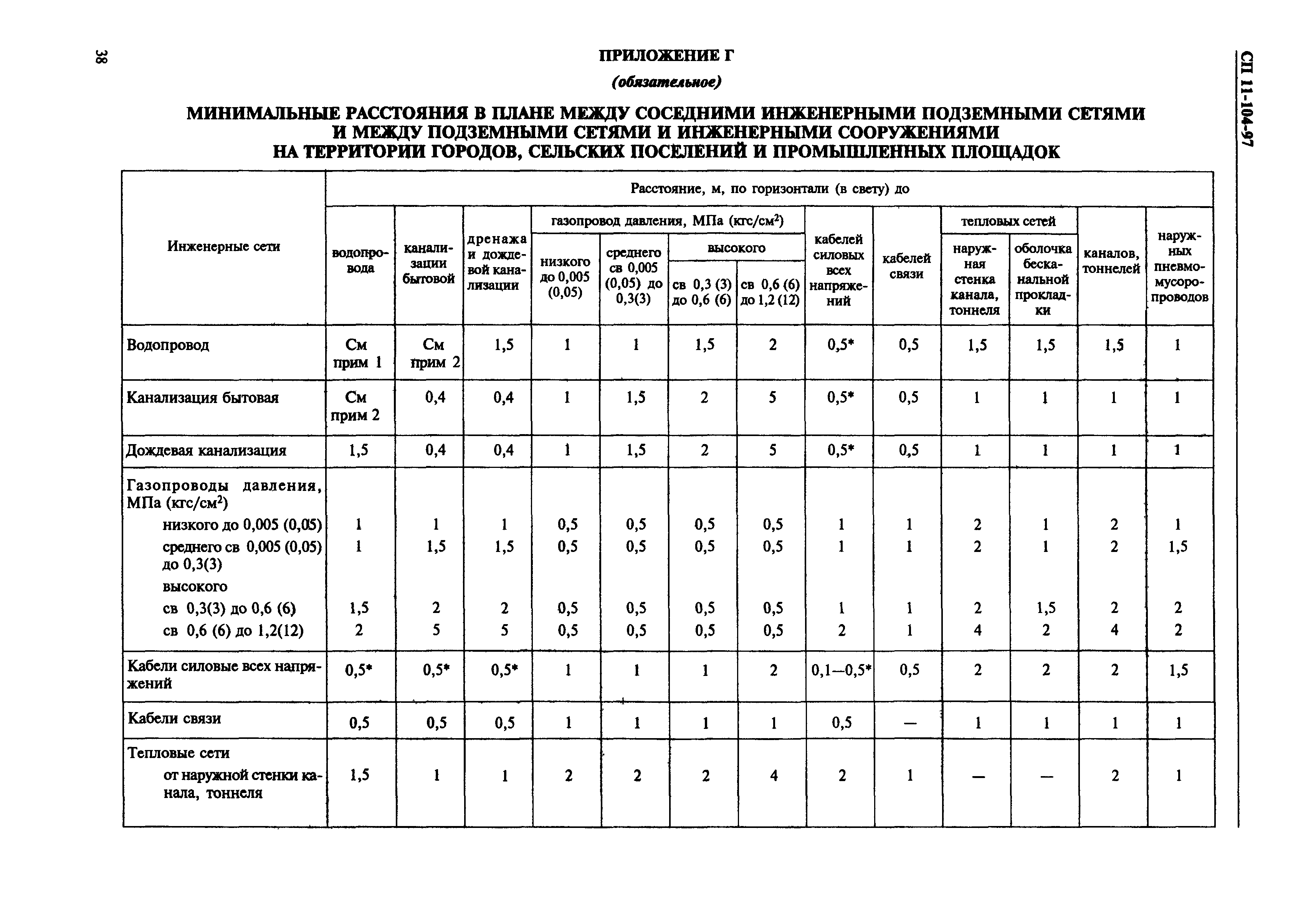
Согласно пункту 4 Типовых правил, основополагающим в данной отрасли при обеспечении безопасности тепловых сетей является СНиП 41-02-2003 «Тепловые сети». В этом СНиП есть специальное приложение, в котором можно посмотреть расстояния по горизонтали от трубопровода до зданий и инженерных сооружений.
Приказ № 115 Минэнерго Российской Федерации от апреля 2003 года не разрешает эксплуатацию тепловых энергоустановок в местах, где прокладываются теплопроводы. Это относится ко всем организациям, каким бы статусом они ни были наделены. Запретительные документы были утверждены арбитражным судом.
Это относится ко всем организациям, каким бы статусом они ни были наделены. Запретительные документы были утверждены арбитражным судом.
Таблица удаленности коммуникаций по нормам СНиП и СП
Охранная зона коммуникаций по поставке тепла формируется без наличия единого целостного подхода, который был бы оформлен в действующем законодательстве.
При дистанцировании от линии подачи электроэнергии можно воспользоваться Постановлением Правительства РФ № 160 по электросетевому хозяйству и Земельным кодексом. Последний документ законодательно закрепляет возможность создания особых зон и эксплуатации участков, расположенных в них.
Расстояния от инженерных сетей до деревьев и кустарников согласно СП и СНиП
Список запретов в охранных зонах тепловых сетей
СНиП 41-02-2003, утвержденный как основное руководство к определению расстояний, только устанавливает требования относительно минимальных дистанций. В населенных пунктах требуется выдерживать расстояния до линейных объектов.
Надземная теплотрасса около города
Однако, как и в случае со зданиями и сооружениями любого типа, отступы могут варьироваться в зависимости от климата. При этом учитывается сила ветра, а также температурные перепады. Многое зависит от вида прокладки – надземной или подземной, рельефа местности и плотности застройки.
Прежде всего в нормативных документах указывается, что охранная зона должна быть территорией, на которой соблюдаются запреты на строительство.
Также недопустимо присутствие объектов, представляющих потенциальное препятствие к нормальному функционированию и ремонтным работам.
Учитывается возможность повреждения, аварии, причинения вреда здоровью граждан. Для проведения определенных работ физические лица и организации должны получить письменное согласие владельцев теплосетей.
Дистанции от труб канализации до различных объектов по нормативам СНиП последней редакции
Без него в защитной зоне запрещается:
- загромождение оборудованных подъездов или подходов к объектам теплопоставок, для которых обозначена зона тепловых сетей;
- специально обозначенная зона теплотрассы не может быть местом для размещения АЗС с бензином или газовыми резервуарами, на ней нельзя складировать ни горюче-смазочные материалы, ни химические соединения агрессивного типа;
- зона тепловых сетей не может использоваться даже для временных сооружений вроде стоянок машин и спортплощадок, а тем более для огородов, гаражей и остановок общественного транспорта;
- охранная зона не может предназначаться для свалок, слива едких и горючих жидкостей, сжигания мусора, погрузочных работ, рытья каналов под другие коммуникации, установки ограждений, посадки зеленых насаждений, где кусты и ветки будут представлять потенциальные помехи при проведении ремонта.

Нормы расположения инженерных сетей согласно СП и СНиП
Тонкости расположения тепловых сетей
Получение письменного разрешения для проведения строительных работ, реконструкции зданий и сооружений и даже планировки грунта в целях благоустройства должно осуществляться не позднее, чем за три дня до предполагаемого старта.
Зона тепловых сетей – территория, статус которой необходим для соблюдения любой организацией, а также предприятиями, отвечающими за другие охранные зоны. В частности, электролиний, газопроводов, трамвайных и железнодорожных путей, автострад и шоссе федерального значения.
Трубы подземного теплопровода
Но даже наличие письменного разрешения не может противоречить нормам, регламентированным в СНиП (СП), и Типовым правилам. Согласно данным нормативным документам нельзя загромождать доступ к наружным теплосетям и объектам обслуживания. Нормы удаленности в каждую сторону предусмотрены в целях охраны теплопровода.
Как определяется охранная зона
Стандартное требование к соблюдению расстояний по нормам СНиП 41-02-2003 предусматривает отступ от края тепловой сети в каждую из сторон по месту прокладки не менее 3 метров. Однако у региональных властей есть возможность самостоятельно решать, сколько метров в каждую сторону будет составлять отступ в конкретном случае.
Однако у региональных властей есть возможность самостоятельно решать, сколько метров в каждую сторону будет составлять отступ в конкретном случае.
Надземная теплотрасса
Определяющими факторами могут стать климатические условия, тип прокладки тепловой сети (воздушный или подземный), тип грунта, диаметр труб, напор и температура подачи.
При этом в каждую сторону можно только увеличивать отступ. Однако решение, сколько метров составляет охранная зона, не может быть меньше, чем указанное в нормативных документах.
Охранная зона подземной теплотрассы, согласно последнему Федеральному Закону, принятому по этой тематике, регистрируется в ЕГРН (Едином Государственном реестре недвижимости). Сведения о том, где на конкретной территории находится зона тепловых сетей, можно получить не только в управлении строительства и архитектуры или у владельцев теплосетей, но и в ЕГРН, куда обязаны подавать сведения местные органы самоуправления.
Трубы для подземной прокладки
В нормативном акте об обязательной регистрации, где зона теплотрассы регистрируется как земельная собственность, обязательно указывается, сколько метров составляют официально зарегистрированные ограничения от тепловых сетей.
На территории санитарной зоны тепловых сетей нельзя без разрешения посадить даже кустарник. Над воздушным трубопроводом не должны находиться толстые ветки деревьев, способные привести к аварии трубопровода.
Если нарушается охранная зона теплоподающей сети и вспомогательных сооружений, владелец может обратиться в суд. Если это нарушение привело к поломке или аварии, стоимость ущерба взыскивается с нарушителя запретной границы.
Подземная теплотрасса в бетонном канале
Правовые аспекты и тепловые сети
Последние решения арбитражного суда, редакции строительных норм и правил, постановлений, принятых для рассмотрения новых видов исков о нарушении законных прав владельца земельной собственности, ужесточили ответственность нарушителей по категории судебных дел «Правонарушение без лишения владения».
Трубы для наружной прокладки теплотрассы
Теперь суд может рассматривать такие дела и устанавливать административное наказание на основании зарегистрированной земельной собственности в теплопроводной охранной зоне.
Там документально зафиксировано, сколько официально метров должен составлять отступ в каждую сторону от тепловой сети.
При рассмотрении судебного иска против физического лица или организации при нарушении прав владельцев теплосети, решение может быть не только о прекращении строительства, реконструкции, земельного благоустройства.
Зона ремонта теплосети
Суд может постановить устранить уже возведенные постройки. Причем в этом случае срок исковой давности не имеет значения. Решение может обязывать снести или перенести объекты, находящиеся в охранной зоне, если документально подтвержден факт нарушения имущественных прав, норм градостроения и строительства.
СНиП 2.
 04.07-86 (Приложение 23) / Pozhproekt.ru
04.07-86 (Приложение 23) / Pozhproekt.ruПРИЛОЖЕНИЕ 23* (Обязательное)
РАСЧЕТНЫЙ РАСХОД ВОДЫ ДЛЯ ПОДПИТКИ ВОДЯНЫХ ТЕПЛОВЫХ СЕТЕЙ, ЧИСЛО И ЕМКОСТЬ БАКОВ-АККУМУЛЯТОРОВ И БАКОВ ЗАПАСА ПОДПИТОЧНОЙ ВОДЫ И ТРЕБОВАНИЯ ПО ИХ УСТАНОВКЕ
1. Расчетный расход воды, м3/ч, для подпитки тепловых сетей следует принимать:
а) в закрытых системах теплоснабжения — численно равным 0,75% фактического объема воды в трубопроводах тепловых сетей и присоединенных к ним системах отопления и вентиляции зданий. При этом для участков тепловых сетей длиной более 5 км от источников теплоты без распределения теплоты расчетный расход воды следует принимать равным 0,5% объема воды в этих трубопроводах;
б) в открытых системах теплоснабжения — равным расчетному среднему расходу воды на горячее водоснабжение с коэффициентом 1,2 плюс 0,75% фактического объема воды в трубопроводах тепловых сетей и присоединенных к ним системах отопления, вентиляции и горячего водоснабжения зданий. При этом для участков тепловых сетей длиной более 5 км от источников теплоты без распределения теплоты расчетный расход воды следует принимать равным 0,5% объема воды в этих трубопроводах;
При этом для участков тепловых сетей длиной более 5 км от источников теплоты без распределения теплоты расчетный расход воды следует принимать равным 0,5% объема воды в этих трубопроводах;
в) для отдельных тепловых сетей горячего водоснабжения при наличии баков-аккумуляторов — равным расчетному среднему расходу воды на горячее водоснабжение с коэффициентом 1,2; при отсутствии баков — по максимальному расходу воды на горячее водоснабжение плюс (в обоих случаях) 0,75% фактического объема воды в трубопроводах сетей и присоединенных к ним системах горячего водоснабжения зданий.
2. Объем воды в системах теплоснабжения при отсутствии данных по фактическим объемам воды допускается принимать равным 65 м3 на 1 МВт расчетного теплового потока при закрытой системе теплоснабжения 70 м3 на 1 МВт — при открытой системе и 30 м3 на 1 МВт — при отдельных сетях горячего водоснабжения.
3. Для открытых и закрытых систем теплоснабжения должна предусматриваться дополнительно аварийная подпитка химически не обработанной и не деаэрированной водой, расход которой принимается в количестве 2% объема воды в трубопроводах тепловых сетей и присоединенных к ним системах отопления, вентиляции и в системах горячего водоснабжения для открытых систем теплоснабжения.
При наличии нескольких отдельных тепловых сетей, отходящих от коллектора теплоисточника, аварийную подпитку допускается определять только для одной наибольшей по объему тепловой сети.
Для открытых систем теплоснабжения аварийная подпитка должна обеспечиваться только из систем хозяйственно-питьевого водоснабжения.
4. Для открытых систем теплоснабжения, а также при отдельных тепловых сетях на горячее водоснабжение должны предусматриваться баки-аккумуляторы химически обработанной и деаэрированной подпиточной воды, расчетной емкостью равной десятикратной величине среднего расхода воды на горячее водоснабжение.
В закрытых системах теплоснабжения на источниках теплоты мощностью 100 МВт и более следует предусматривать установку баков запаса химически обработанной и деаэрированной подпиточной воды емкостью 3% объема воды в системе теплоснабжения. схема включения баков запаса должна обеспечивать непрерывнее обновление воды в баках. Количество баков независимо от системы теплоснабжения принимается не менее двух по 50% рабочего объема.
5. Размещение баков-аккумуляторов горячей воды возможно как на источнике теплоты, так и в районах теплопотребления. При этом на источнике теплоты должны предусматриваться баки-аккумуляторы емкостью не менее 25% общей расчетной емкости баков. На территории источников теплоты установку баков-аккумуляторов следует предусматривать по нормам Минтопэнерго России.
6. Внутренняя поверхность баков должна быть защищена от коррозии, а вода в них—от аэрации.
7. Группа баков должна быть ограждена валом высотой не менее 0,5 м. Обвалованная территория должна вмещать объем наибольшего бака и иметь отвод воды в канализацию.
8. Устанавливать баки-аккумуляторы горячей воды в жилых кварталах не допускается. Расстояние от баков-аккумуляторов горячей воды до границы жилых кварталов должно быть не менее 30 м. При этом на грунтах I типа просадочности расстояние, кроме того, должно быть не менее 1,5 толщины слоя просадочного грунта.
При размещении баков-аккумуляторов вне территории источников теплоты следует предусматривать их ограждение высотой не менее 2,5 м для исключения доступа посторонних лиц к бакам.
SNP MP приветствует роль крупнейшей в Великобритании схемы централизованного теплоснабжения с использованием возобновляемых источников энергии в борьбе с топливной бедностью
Член парламента Дейрдре Брок демонстрирует схему централизованного теплоснабжения Уиндфорда в Глазго.Депутат SNP Дейдре Брок посетила инновационную схему централизованного теплоснабжения Wyndford в Глазго, чтобы узнать больше о том, как сделать старые дома более энергоэффективными и соответствовать повестке дня Великобритании по низкому уровню выбросов углерода .
Визит был организован Ассоциацией децентрализованной энергетики (ADE), и Брок (член парламента от Лейта и северного Эдинбурга) продемонстрировал, как работает централизованное теплоснабжение и как эта схема помогает улучшить качество жизни жителей и решить проблему топливной бедности в длительный срок.
Схема Wyndford является домом для одной из крупнейших в Великобритании схем модернизации централизованного теплоснабжения, обеспечивающей по требованию низкоуглеродное отопление и горячую воду для более чем 1800 домов. Система была установлена SSE в партнерстве с шотландской жилищной ассоциацией.
Система была установлена SSE в партнерстве с шотландской жилищной ассоциацией.
В недавнем отчете, опубликованном SSE, показано, что схема позволила сократить выбросы CO2 на 62% с момента ее установки, а также убедительные доказательства того, что жизнь людей значительно улучшилась, повысился комфорт, были созданы рабочие места и увеличилась экономическая ценность.
Комментируя экскурсию по энергоцентру, Брок сказал: «Централизованное теплоснабжение — это грандиозная идея — судя по тому, что я видел, для создания более теплых домов и повышения энергоэффективности. Это, конечно, делает его более экологически чистым, так что и планета в выигрыше.
«Возможно, это будет следующим шагом, который Шотландия должна предпринять, чтобы одновременно избавиться от топливной бедности и экологически безопасного отопления.
«Я немного узнал о том, как работает система во время этого визита, и она была модернизирована, поэтому повсюду есть возможности, если мы сможем реализовать схемы. ”
”
См. Также: —
Скотт-Правительство должно срочно изменить приоритеты после неспособности искоренить топливную бедность
Energy charity urges Scot-Govt to urgently re-set priorities after failing to eradicate fuel-poverty
Картирование QTL для термостойкости риса с высоким разрешением с использованием массива SNP 5K | Рис
Begum T, Reuter R, Schöffl F (2013) Сверхэкспрессия AtHsfB4 вызывает специфические эффекты на развитие корней Arabidopsis.Механический Дев 130 (1): 54–60. DOI: 10.1016 / j.mod.2012.05.008
CAS Статья PubMed Google ученый
Broman KW, Wu H, Sen Ś, Churchill GA (2003) R / qtl: Картирование QTL в экспериментальных скрещиваниях. Биоинформатика 19 (7): 889–890
Биоинформатика 19 (7): 889–890
CAS Статья PubMed Google ученый
Buu BC, Ha PTT, Tam BP, Nhien TT, Hieu NV, Phuoc NT, Minh LT, Giang LH, Lang NT (2014) Локусы количественных признаков, связанные с термостойкостью риса ( Oryza sativa L .) Порода растений Биотех 2: 14–24
Статья Google ученый
Cao L, Zhao J, Zhan X, Li D, He L, Cheng S (2003) Картирование QTL для термостойкости и корреляции между термостойкостью и скоростью фотосинтеза в рисе. Chin J Rice Sci 17: 223–227
CAS Google ученый
Carriger S, Vallee D (2007) Больше урожая на каплю. Рис сегодня 6: 10–13
Рис сегодня 6: 10–13
Google ученый
Chang-Lan Z, Ying-Hui X, Chun-Ming W, Ling J, Hu-Qu Z, Jian-Min W. (2005) Отображение QTL для термостойкости на стадии заполнения зерна в рис.Rice Sci 12 (1): 33–38
Google ученый
Chen M, Presting G, Barbazuk WB, Goicoechea JL, Blackmon B, Fang G, Kim H, Frisch D, Yu Y, Sun S, Higingbottom S, Phimphilai J, Phimphilai D, Thurmond S, Gaudette B, Li П, Лю Дж., Хэтфилд Дж., Мэйн Д., Фаррар К., Хендерсон К., Барнетт Л., Коста Р., Уильямс Б., Уолзер С., Аткинс М., Холл С, Будиман М. А., Томкинс Дж. П., Луо М., Бэнкрофт И., Сальсе Дж., Регад Ф., Мохапатра Т., Сингх Н.К., Тьяги А.К., Содерлунд С., Дин Р.А., Винг Р.А. (2002) Интегрированная физическая и генетическая карта генома риса. Растительная клетка 14 (3): 537–545
Растительная клетка 14 (3): 537–545
Статья PubMed PubMed Central Google ученый
Chen Q, Yu S, Li C, Mou T (2008) Идентификация QTL для термостойкости риса на стадии цветения. Sci Agric Sin 41: 315–321
CAS Google ученый
Cheng L, Wang J, Uzokwe V, Meng L, Wang Y, Sun Y, Zhu L, Xu J, Li Z (2012) Генетический анализ устойчивости к холоду на стадии проростков и устойчивости к теплу при цветении риса ( Oryza sativa L.) J Integ Agriculture 11: 359–367
Статья Google ученый
Дойл Дж. Дж., Дойл Дж. Л. (1990) Выделение растительной ДНК из свежей ткани. Фокус 12: 13–15
Фокус 12: 13–15
Google ученый
Fernandez GCJ (1992) Эффективные критерии отбора для оценки устойчивости растений к стрессу. В: Кус Э.Г. (ред.) Адаптация температуры пищевых культур и водного стресса. Материалы 4-го Международного симпозиума Азиатского центра овощей и исследований и развития, Шантана, Тайвань, стр. 257–270
Google ученый
Фишер Р.А., Маурер Р. (1978) Засухоустойчивость сортов яровой пшеницы.I. Ответы на урожайность зерна. Aust J Agric Res 29: 892–912
Google ученый
Гарг А.К., Ким Дж.К., Оуэнс Т.Г., Ранвала А.П., Чой Ю.Д., Кочиан Л.В. , Ву Р.Дж. (2002) Накопление трегалозы в растениях риса обеспечивает высокий уровень толерантности к различным абиотическим стрессам. PNAS 99 (10): 15898–15903. DOI: 10.1073 / pnas.252637799
, Ву Р.Дж. (2002) Накопление трегалозы в растениях риса обеспечивает высокий уровень толерантности к различным абиотическим стрессам. PNAS 99 (10): 15898–15903. DOI: 10.1073 / pnas.252637799
CAS Статья PubMed PubMed Central Google ученый
Giorno F, Wolters-Arts M, Mariani C, Ivo R (2013) Обеспечение воспроизводства при высоких температурах: реакция на тепловой стресс во время развития пыльников и пыльцы.Растения 2: 489–506. DOI: 10.3390 / растения2030489
Артикул PubMed PubMed Central Google ученый
Guo J, Wu J, Ji Q, Wang C, Luo L, Yuan Y, Wang Y, Wang J (2008) Полногеномный анализ семейств факторов транскрипции теплового шока у риса и Arabidopsis. Дж. Генет Геномика 35 (2): 105–118. DOI: 10.1016 / S1673-8527 (08) 60016-8
Дж. Генет Геномика 35 (2): 105–118. DOI: 10.1016 / S1673-8527 (08) 60016-8
CAS Статья PubMed Google ученый
IPCC (2007) Резюме для политиков.В: Соломон С., Цинь Д., Мэннинг М., Чен З., Маркиз М., Аверит К. Б., Тигнор М., Миллер Х. Л. (ред.) Изменение климата 2007: основы физической науки. Вклад рабочей группы I в четвертый оценочный доклад межправительственной группы экспертов по изменению климата. Издательство Кембриджского университета, Кембридж, стр. 1–18
Google ученый
Jagadish SVK, Cairns J, Lafitte R, Wheeler TR, Price AH, Craufurd PQ (2010b) Генетический анализ термостойкости при цветении риса.Crop Sci 50: 1633–1641
CAS Статья Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Jagadish SVK, Craufurd PQ, Wheeler TR (2007) Высокотемпературный стресс и фертильность колосков в рисе ( Oryza sativa L. ) J Exp Bot 58 (7): 1627–1635
CAS Статья PubMed Google ученый
Jagadish SVK, Muthurajan R, Oane R, Wheeler TR, Heuer S, Bennett J, Craufurd PQ (2010a) Физиологические и протеомные подходы к решению проблемы термостойкости во время цветения риса ( Oryza sativa L.) J Exp Bot 61: 143–156. DOI: 10.1093 / jxb / erp289
CAS Статья PubMed Google ученый
Jagadish SVK, Craufurd PQ, Wheeler TR (2008) Фенотипирование родителей картируемых популяций риса ( Oryza sativa L.) на предмет термостойкости во время цветения. Crop Sci 48: 1140–1146
Crop Sci 48: 1140–1146
Статья Google ученый
Kaplan F, Kopka J, Haskell DW, Zhao W., Schiller KC, Gatzke N, Sung DY, Guy CL (2004) Изучение температурно-стрессового метаболома Arabidopsis.Физиология растений 136 (4): 4159–4168
CAS Статья PubMed PubMed Central Google ученый
Kong D, Li M, Dong Z, Ji H, Li X (2015) Идентификация TaWD40D, белка, содержащего повтор WD40 пшеницы, который связан с устойчивостью растений к абиотическим стрессам. Plant Cell Rep 3: 395–410
Статья Google ученый
Kumar V, Singh A, Mithra SVA, Krishnamurthy SL, Parida SK, Jain S, Tiwari KK, Kumar P, Rao AR, Sharma SK, Khurana JP, Singh NK, Mohapatra T (2015) Картирование ассоциаций в масштабе всего генома солеустойчивости риса ( Oryza sativa L).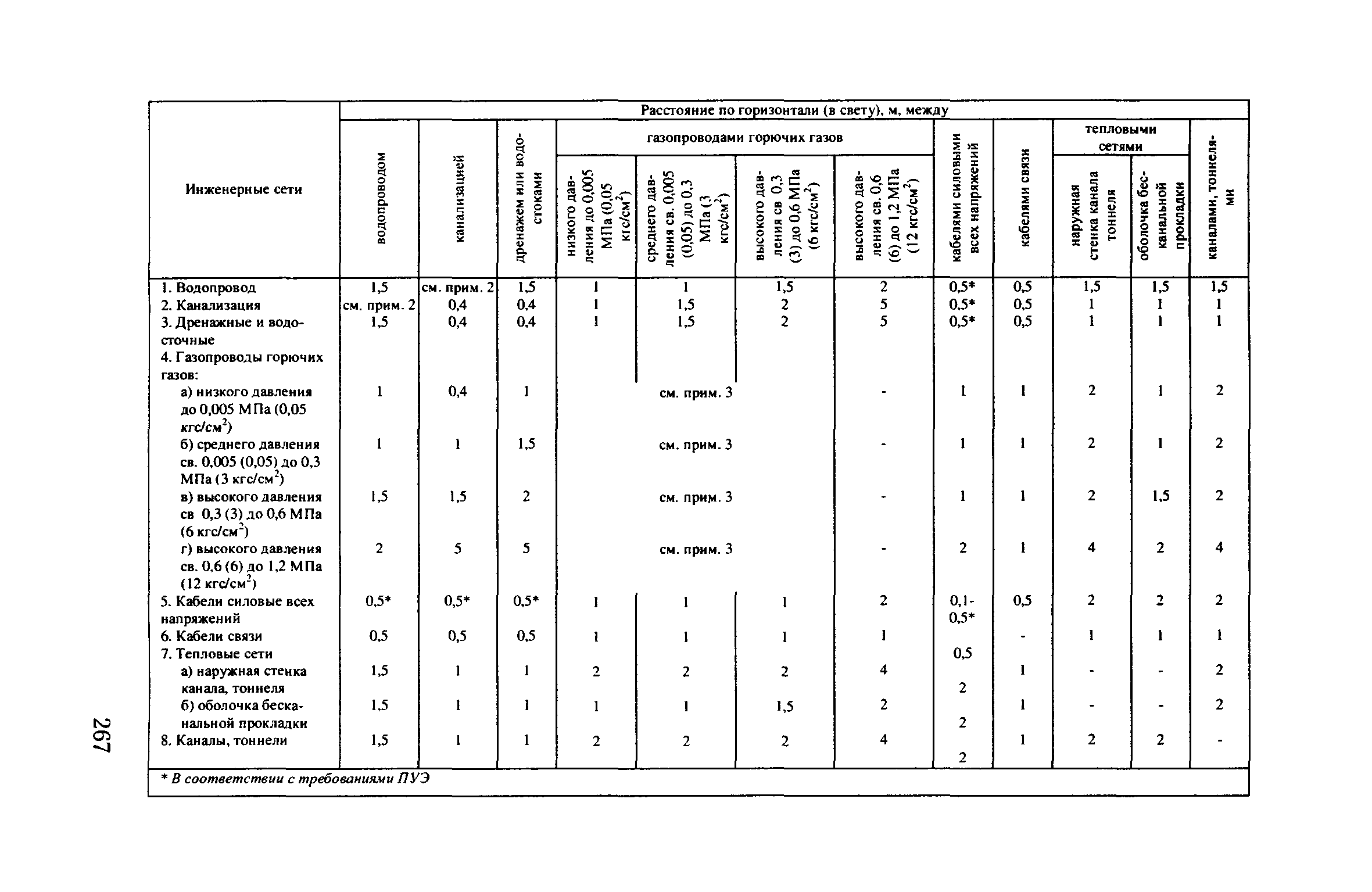 ДНК Res 22 (2): 133–145. DOI: 10.1093 / dnares / dsu046
ДНК Res 22 (2): 133–145. DOI: 10.1093 / dnares / dsu046
CAS Статья PubMed PubMed Central Google ученый
Li H, Yuan Z, Vizcay-Barrena G, Yang C, Liang W, Zong J, Wilson ZA, Zhang D (2011a) ПОСТОЯННАЯ ТАПЕТАЛЬНАЯ КЛЕТКА1 кодирует белок PHD-finger, который необходим для гибели тапетальных клеток и пыльцы развитие в рисе. Физиология растений 156 (2): 615–630. DOI: 10.1104 / стр.111.175760
CAS Статья PubMed PubMed Central Google ученый
Li HW, Zang BS, Deng XW, Wang XP (2011b) Сверхэкспрессия гена трегалозо-6-фосфатсинтазы OsTPS1 повышает устойчивость риса к абиотическому стрессу.Planta 234 (5): 1007–1018. DOI: 10.1007 / s00425-011-1458-0
DOI: 10.1007 / s00425-011-1458-0
CAS Статья PubMed Google ученый
Li WL, Liu Y, Douglas CJ (2016) Роль гликозилтрансфераз в образовании примексина стенки пыльцы и формировании паттерна экзин. Plant Physiol. 173; (1): 167–82.
Li N, Zhang DS, Liu HS, Yin CS, Li XX, Liang WQ, Yuan Z, Xu B, Chu HW, Wang J, Wen TQ, Huang H, Luo D, Ma H, Zhang DB (2006 ) Ген задержки дегенерации тапетума риса необходим для деградации тапетума и развития пыльников.Растительная клетка 18: 2999–3014
CAS Статья PubMed PubMed Central Google ученый
Liu C, Qi X, Zhao Q, Yu J (2013) Характеристика и функциональный анализ специфичного для пыльцы картофеля белка SBgLR, ассоциированного с микротрубочками, в табаке. PLoS One 8 (3): e60543. DOI: 10.1371 / journal.pone.0060543
PLoS One 8 (3): e60543. DOI: 10.1371 / journal.pone.0060543
CAS Статья PubMed PubMed Central Google ученый
Liu JX, Liao DQ, Oane R, Estenor L, Yang XE, Li ZC, Bennett J (2006) Генетические вариации чувствительности расхождения пыльников к стрессу засухи у риса.Field Crop Res 97: 87–100
Статья Google ученый
Лобелл Д.Б., Шленкер В., Коста-Робертс Дж. (2011) Климатические тенденции и мировое растениеводство с 1980 года. Наука 333: 616–620
CAS Статья PubMed Google ученый
Lohmann C, Eggers-Schumacher G, Wunderlich M, Schöffl F (2004) Два разных фактора транскрипции теплового шока регулируют немедленную раннюю экспрессию стрессовых генов у Arabidopsis.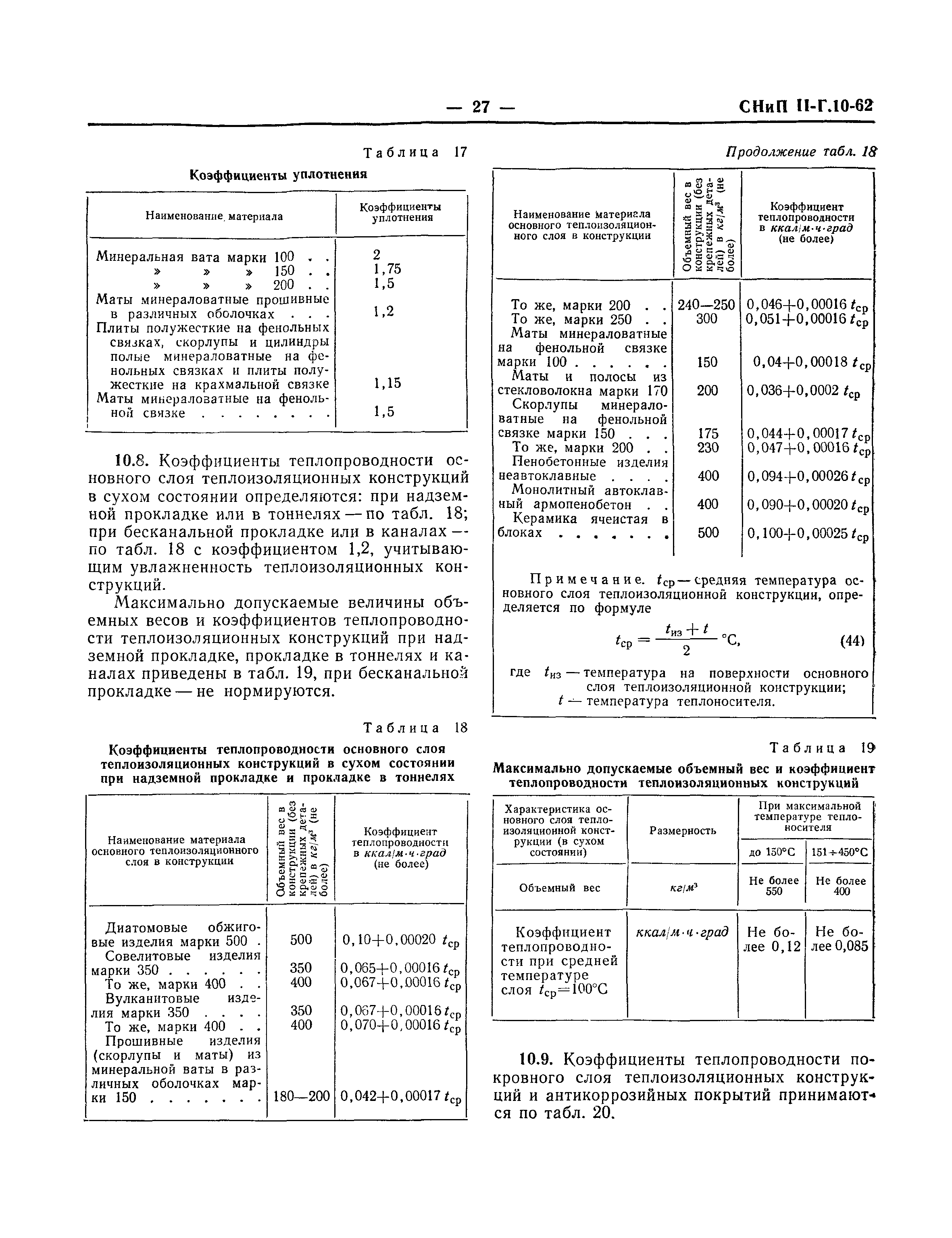 Mol Gen Genomics 271 (1): 11–21
Mol Gen Genomics 271 (1): 11–21
CAS Статья Google ученый
Mansueto L, Fuentes RR, Borja FN, Detras J, Abriol-santos M, ChebotarovD,… Александров Н. (2017) Обновление базы данных Rice SNP-seek: новые SNP, InDels и запросы. Нуклеиновые кислоты Res 45: D1075-D1081.
Мацуи Т., Омаса К., Хори Т. (1997) Высокая температура, вызванная стерильностью колосков риса японской во время цветения, в зависимости от влажности воздуха и условий скорости ветра.Japan Journal of Crop Science 66: 449–455
Статья Google ученый
Мацуи Т., Омаса К., Хори Т. (1999) Быстрое набухание пыльцевых зерен в ответ на раскрытие цветочка раскрывает локул в рисе. Наука о растениеводстве 2 (3): 196–199
Наука о растениеводстве 2 (3): 196–199
Статья Google ученый
Мацуи Т., Омаса К. (2002) Сорта риса ( Oryza sativa L.), устойчивые к высокой температуре во время цветения: характеристики пыльников.Ann Bot 89 (6): 683–637
Статья PubMed PubMed Central Google ученый
Meng L, Li H, Zhang L, Wang J (2015) QTL IciMapping: интегрированное программное обеспечение для построения карты генетического сцепления и количественного картирования локусов признаков в популяции с двумя родителями. Урожай J 3: 265–279. DOI: 10.1016 / j.cj.2015.01.001
Артикул Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Miranda JA, Avonce N, Suárez R, Thevelein JM, Van Dijck P, Iturriaga G (2007) Бифункциональный фермент TPS-TPP из дрожжей придает устойчивость к множественным и экстремальным условиям абиотического стресса у трансгенного арабидопсиса.Planta 226 (6): 1411–1421
CAS Статья PubMed Google ученый
Мишра А.К., Пураник С., Прасад М. (2012) Структура и регуляторные сети белка WD40 в растениях. J Plant Biochem Biotechnol 21: 32–39
CAS Статья Google ученый
Мохапатра Т., Робин С., Сарла Н., Шешашай М., Сингх А.К., Сингх К., Сингх Н.К., Амитха Митра С.В., Шарма Р.П. (2014) EMS-индуцированные мутанты сорта риса нагина22: создание и характеристика.Proc Indian National Science Academy 80: 163–172
Статья Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Moon S, Kim SR, Zhao G, Yi J, Yoo Y, Jin P, Lee SW, Jung KH, Zhang D, An G (2013) Rice GLYCOSYLTRANSFERASE1 кодирует гликозилтрансферазу, необходимую для образования стенки пыльцы. Физиология растений 161 (2): 663–675. DOI: 10.1104 / стр.112.210948
CAS Статья PubMed Google ученый
Накагава Х., Хори Т., Мацуи Т. (2002) Влияние изменения климата на производство риса и адаптивные технологии.В: Mew TW, Brar DS, Peng S, Dawe D, Hardy B (eds) Наука о рисе: инновации и влияние на средства к существованию Китай: Международный научно-исследовательский институт риса, стр. 635–657
Google ученый
Nuccio ML, Wu J, Mowers R, Zhou HP, Meghji M, Primavesi LF, Paul MJ, Chen X, Gao Y, Haque E, Basu SS, Lagrimini LM (2015) Экспрессия трегалозо-6-фосфатфосфатазы в початках кукурузы повышает урожайность в условиях полива и засухи. Нат Биотех 33: 862–869.DOI: 10.1038 / nbt.3277
Нат Биотех 33: 862–869.DOI: 10.1038 / nbt.3277
CAS Статья Google ученый
Pandit A, Rai V, Bal S, Sinha S, Kumar V, Chauhan M, Gautam RK, Singh R, Sharma PC, Singh AK, Gaikwad K, Sharma TR, Mohapatra T, Singh NK (2010) Объединение QTL картирование и профилирование транскриптомов объемных RIL для идентификации функционального полиморфизма генов солеустойчивости у риса ( Oryza sativa L.) Mol Gen Genomics 284 (2): 121–136.DOI: 10.1007 / s00438-010-0551-6
CAS Статья Google ученый
Poli Y, Basava RK, Panigrahy M, Vinukonda VP, Dokula NR, Voleti SR, Desiraju S, Neelamraju S (2013) Характеристика мутанта риса Nagina22 по термостойкости и картографирование признаков урожайности. Рис 6:36
Рис 6:36
Артикул PubMed PubMed Central Google ученый
Prakash C, Mithra SVA, Singh PK, Mohapatra T, Singh NK (2016) Раскрытие молекулярных основ управления окислительным стрессом в засухоустойчивом генотипе риса Nagina22.BMC Genomics 17: 774. DOI: 10.1186 / s12864-016-3131-2
Артикул PubMed PubMed Central Google ученый
Prasad PVV, Boote KJ, Jr LHA, Sheehy JE, Thomas JMG (2006) Различия между видами, экотипами и сортами в плодовитости колосков и индексе урожая риса в ответ на высокотемпературный стресс. Field Crop Res 95: 398–411
Статья Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Qi Y, Liu Q, Zhang L, Mao B, Yan D, Jin Q, He Z (2014) Точное картирование и анализ генов-кандидатов нового гена термочувствительной мужской генной стерильности tms9-1 в рисе.Theor Appl Genet 127 (5): 1173–1182. DOI: 10.1007 / s00122-014-2289-8
CAS Статья PubMed Google ученый
Raman A, Verulkar SB, Mandal N, Variar M, Shukla VD, Dwivedi JL, Singh BN, Singh ON, Swain P, Mall AK, Robin S, Chandrababu R, Jain A, Ram T, Hittalmani S, Haefele S, Piepho HP, Kumar A (2012) Индекс засухи для выбора линий высокоурожайного риса в условиях разной степени стресса от засухи.Рис 5:31. DOI: 10.1186 / 1939-8433-5-31
Артикул PubMed Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Сарвар М., Авеси Г.М. (1985) Оценка зародышевой плазмы риса на устойчивость к высоким температурам. Пакистан, журнал J Agric Res 6 (3): 162–164
Google ученый
Сато К., Инаба К., Тосава М. (1973) Высокотемпературное повреждение созревания растений риса. Влияние высокотемпературной обработки на разных стадиях развития метелки на созревание.Труды Японского общества растениеводства 42: 207–213
CAS Статья Google ученый
Смертенко А.П., Чанг Х.Й., Вагнер В., Калорити Д., Феник С., Сонобе С., Ллойд С., Хаузер М.Т., Хасси П.Дж. (2004) The Arabidopsis Microtubule-Associated Protein AtMAP65 -1: молекулярный анализ его активность связывания микротрубочек. Растительная клетка 16 (8): 2035–2047
Растительная клетка 16 (8): 2035–2047
CAS Статья PubMed PubMed Central Google ученый
Szalonek M, Sierpien B, Rymaszewski W, Gieczewska K, Garstka M, Lichocka M, Sass L, Paul K, Vass I, Vankova R, Dobrev P, SzczesnyP MW, Krusiewicz D, Hen-Zzelczyk , Konopka-Postupolska D (2015) Аннексин картофеля STANN1 способствует устойчивости к засухе и снижает световой стресс у трансгенных растений Solanum tuberosum L.Растения. PLoS One 10 (7): e0132683. DOI: 10.1371 / journal.pone.0132683
Артикул PubMed PubMed Central Google ученый
Tazib T, Kobayashi Y, Koyama H, Matsui T (2015) QTL-анализ длины пыльника и растяжения во время цветения как признаков устойчивости риса к экстремальным температурам ( Oryza sativa L. ) Euphytica 203: 629– 642
) Euphytica 203: 629– 642
Артикул Google ученый
Tiwari S, Krishnamurthy SL, Kumar V, Singh B, Rao A, Mithra SVA, Rai V, Singh AK, Singh NK (2016) Отображение QTL для толерантности к соли в рисе ( Oryza sativa L.) с помощью массового сегрегантного анализа рекомбинантных инбредных линий с использованием чипа SNP 50K. PLoS One 11 (4): e0153610. DOI: 10.1371 / journal.pone.0153610
Артикул PubMed PubMed Central Google ученый
Организация Объединенных Наций, Департамент по экономическим и социальным вопросам, Отдел народонаселения (2011 г.) Перспективы народонаселения мира: издание 2010 г., Том I: Комплексные таблицы. ST / ESA / SER.A / 313
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Xiao Y, Pan Y, Luo L, Zhang G, Deng H, Dai L, Liu X, Tang W, Chen L, Wang GL (2011a) Локусы количественных признаков, связанные с Завязавшиеся семена риса ( Oryza sativa L.) Euphytica 178: 331–338. DOI: 10.1007 / s10681-010-0300-2
Артикул Google ученый
Xiao YH, Pan Y, Luo LH, Deng HB, Zhang GL, Tang WB, Chen LY (2011b) Локусы количественных признаков, связанные с фертильностью пыльцы в условиях высокотемпературного стресса на стадии цветения риса ( Oryza sativa L ) . Rice Sci 18 (2): 1–7
Google ученый
Xu L, Tang Y, Gao S, Su S, Hong L, Wang W, Fang Z, Li X, Ma J, Quan W, Sun H, Li X, Wang Y, Liao X, Gao J, Zhang F, Li L, Zhao C (2016) Комплексный анализ семейства генов аннексина в пшенице. BMC Genomics 17: 415. DOI: 10.1186 / s12864-016-2750-у
BMC Genomics 17: 415. DOI: 10.1186 / s12864-016-2750-у
Артикул PubMed PubMed Central Google ученый
Янг С., Бискай-Баррена С., Коннер К., Уилсон З.А. (2007) МУЖСКАЯ СТЕРИЛЬНОСТЬ1 требуется для развития тапеталов и биосинтеза пыльцевой стенки. Растительная клетка 19 (11): 3530–3548
CAS Статья PubMed PubMed Central Google ученый
Ye C, Argayoso MA, Redoña ED, Sierra SL, Laza MA, Dilla CJ, Mo Y, Thomson MJ, Chin J, Delaviña CB, Diaz GQ, Hernandez JE (2012) Картирование QTL для термостойкости на стадии цветения в рисе с использованием маркеров SNP.Порода растений 131 (1): 33–41
CAS Статья Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Ye C, Tenorio FA, Argayoso MA, Laza MA, Koh HJ, Redoña ED, Jagadish KS, Gregorio GB (2015) Идентификация и подтверждение локусов количественных признаков, связанных с термостойкостью на стадии цветения в различных популяциях риса. BMC Genet 16:41
Статья PubMed PubMed Central Google ученый
Zhang C, Zhang F (2015) Многофункциональность белков WD40 в целостности генома и развитии клеточного цикла.J Genomics 3: 40–50
Статья PubMed PubMed Central Google ученый
Zhang G, Chen L, Xiao G, Xiao Y, Chen X, Zhang S (2009) Объемный сегрегантный анализ для определения QTL, связанного с термостойкостью риса, с использованием маркеров SSR. Сельское хозяйство Китая 8: 482–487
CAS Статья Google ученый
 org/ScholarlyArticle»>
org/ScholarlyArticle»>Zhang T, Yang L, Jiang K, Huang M, Sun Q, Chen W, Zheng J (2008) Отображение QTL для термостойкости периода кисточки риса.Mol Plant Breed 6: 867–873
CAS Google ученый
Zhao L, Lei J, Huang Y, Zhu S, Chen H, Huang R, Peng Z, Tu Q, Shen X, Yan S (2016) Картирование локусов количественных признаков термостойкости при цветении риса с использованием сегмента хромосомы строки подстановки. Селекционная наука Предварительный просмотр . DOI: 10.1270 / jsbbs.15084.
Zhu J, Wu X, Yuan S, Qian D, Nan Q, An L, Xiang Y (2014) Annexin5 играет жизненно важную роль в развитии пыльцы Arabidopsis через Ca 2+ -зависимый перенос мембран.PLoS One 9 (7): e102407
Артикул PubMed PubMed Central Google ученый
Ответы на ваши вопросы: Как отапливать дом без газа?
Клэр Даймонд
BBC Scotland News
Мы спросили читателей новостного веб-сайта BBC Scotland, что они хотели бы знать об изменении климата. Вот ответы на некоторые из присланных вопросов.
Вот ответы на некоторые из присланных вопросов.
Как будет отапливаться новое жилье без газа? — Ян Рид
Около 80% домов в Шотландии используют центральное газовое отопление, но новые правила будут означать, что с 2024 года новые дома не должны подключаться к газовой сети.
Вместо этого они будут полагаться на возобновляемые источники энергии или альтернативы с низким содержанием углерода.
image copyrightGetty Imagesimage caption Воздушные тепловые насосы — это низкоуглеродный способ отопления домов.Сети централизованного теплоснабжения могут быть вариантом.
Они включают создание горячей воды путем сжигания биомассы, например древесины, которая затем перекачивается по сети в теплоизолированных трубах для отопления и горячего водоснабжения.
Также можно использовать тепловые насосы, которые отбирают тепло из окружающей среды и используют его для обогрева домов.Концепция похожа на холодильник, который отбирает тепло изнутри, чтобы оставаться холодным — в то время как тепловые насосы извлекают тепло извне и доставляют его. проложенные под землей трубы для извлечения тепла из земли.
проложенные под землей трубы для извлечения тепла из земли.
Возобновляемые источники энергии, такие как солнечная, ветровая и гидроэнергетика, также могут способствовать обеспечению электричеством электрических котлов для производства горячей воды и отопления.
Поскольку более 75% электроэнергии, используемой шотландцами, уже поступает из возобновляемых источников, это также может быть более экологичным способом обогрева домов.
Также проводятся испытания, чтобы выяснить, можем ли мы заменить содержимое наших газовых труб водородом.
Кто решает, где размещать точки зарядки автомобилей? — Алан Ланниган
image copyrightGetty ImagesЕсли у вас есть электромобиль, вы можете решить, что хотите, чтобы он был установлен у вас дома.
Компании также могут установить их для сотрудников и клиентов, а местные советы могут также установить общественные точки зарядки.
Департамент транспорта правительства Великобритании заявляет, что в Шотландии в настоящее время имеется более 1500 зарядных устройств, а в Великобритании сейчас больше мест для зарядки, чем заправочных станций.
Как будет заменена ядерная энергия в Шотландии?
image copyrightGetty Imagesimage caption Атомная электростанция Торнесс в Восточном Лотиане — одна из двух атомных электростанций Шотландии.Две атомные электростанции Шотландии — Хантерстон B (которая в настоящее время не производит электроэнергию, пока проводятся проверки безопасности) и Торнесс. Они прекратят вырабатывать электроэнергию в 2023 и 2030 годах соответственно, а правительство Шотландии выступает против строительства новых атомных станций с использованием современных технологий.
Итак, какие возобновляемые и низкоуглеродные источники используются для производства электроэнергии?
Самый крупный источник возобновляемой энергии в Шотландии — морская ветряная электростанция Беатрис — может произвести достаточно энергии для питания 450 000 домов.
Ветряная электростанция Seagreen, которая строится на Ангусе, после завершения строительства сможет обеспечить электроэнергией один миллион домов.
Другие виды возобновляемой электроэнергии от солнечной и гидроэнергетики также могут внести свой вклад.
В будущем приливная энергия также может давать энергию.Но это все еще дорого, и технология требует дальнейшего развития, чтобы поставлять электроэнергию в сеть в коммерческих масштабах.
Достижения в области аккумуляторных технологий могут помочь сгладить некоторые пики и спады производства возобновляемой энергии — так, например, когда меньше ветра для вращения турбин, можно использовать энергию, выработанную ранее.
Правительство Великобритании предлагает снизить пошлины на авиапассажиров. Будет ли это снижение применяться в Шотландии?
image copyrightGetty ImagesAir Passenger Duty (APD) — это налог на пассажиров, вылетающих почти из всех аэропортов Великобритании (хотя аэропорты Хайлендс и острова освобождены от этого налога).
APD был передан Холируду в 2016 году, и правительство Шотландии хочет заменить его новым налогом на выезд из страны.
Компания планировала снизить налог на 50%, прежде чем полностью отменить его, но отказалась от этого предложения после того, как Никола Стерджен объявила чрезвычайную климатическую ситуацию на прошлогодней конференции SNP.
Согласно Financial Times, правительство Великобритании должно пересмотреть APD для всех внутренних рейсов Великобритании в рамках усилий по спасению от проблем с авиакомпанией Flybe. Объявление ожидается в мартовском бюджете.
Если это произойдет, сокращение APD будет применяться в Шотландии до тех пор, пока шотландское правительство, наконец, не возьмет на себя администрирование налога, что, по его словам, будет сделано после того, как будет найдено решение по статусу Хайленда и островов. аэропортов. Затем нужно будет решить, соответствует ли разрез.
Почему путешествие на поезде такое дорогое?
image copyrightGetty Images Прежде всего, строительство и обслуживание — не говоря уже об улучшении — железнодорожной сети — это дорогое удовольствие.
Сменявшие друг друга правительства решили, что расходы на это должны все больше нести пассажиры, а не налогоплательщики.
Таким образом, поезда в Великобритании субсидируются намного меньше, чем в других частях Европы.
Поскольку железные дороги являются естественной монополией, и правительства хотят, чтобы пассажиры платили все большую долю стоимости эксплуатации железных дорог, рост цен на некоторые билеты контролируется государством.
Совсем недавно это увеличение составляло 2.7%.
ScotRail сообщает, что 85% его доходов поступает от тарифов, регулируемых шотландским правительством.
Extinction Rebellion хочет сократить выбросы парниковых газов до нуля к 2025 году. Не могли бы вы спросить их, как этого можно достичь?
image copyrightGetty ImagesОни сказали нам, что хотят, чтобы собрания граждан, которые просят общественность принять решение о политике правительства, сделали трудный выбор.
Какие у вас вопросы о том, что изменение климата означает для Шотландии? Вы можете сообщить нам об этом, заполнив форму ниже.
В некоторых случаях ваш вопрос будет опубликован с отображением вашего имени и местоположения в том виде, в каком вы его предоставили, если вы не укажете иное. Ваши контактные данные никогда не будут опубликованы. Пожалуйста, убедитесь, что вы прочитали условия.
Если вы читаете эту страницу в приложении BBC News, вам нужно будет посетить мобильную версию веб-сайта BBC, чтобы задать свой вопрос по этой теме.
Общегеномное картирование снижения фертильности при тепловом стрессе выявляет QTL, зависящие от стадии развития у Arabidopsis thaliana
ВВЕДЕНИЕ
Одна из основных задач в ближайшие десятилетия — обеспечить рост мирового производства продуктов питания пропорционально увеличению мировое человеческое население.Необходимо поддерживать высокие урожаи, хотя ожидается, что изменение климата приведет к более экстремальным погодным условиям, включая более длительные засухи и более жаркое лето. Селекция растений на устойчивость к абиотическим стрессам, таким как засуха или жара, будет играть решающую роль в решении этой проблемы. Для достижения этой цели крайне важно определить естественный диапазон изменчивости этих черт внутри вида и получить глубокое понимание механистической основы регуляторных процессов. Обе цели могут быть достигнуты путем идентификации ключевых регуляторных генов на больших фенотипических скринингах генетически разнообразных картирующих популяций, полученных естественным или экспериментальным путем.
Для достижения этой цели крайне важно определить естественный диапазон изменчивости этих черт внутри вида и получить глубокое понимание механистической основы регуляторных процессов. Обе цели могут быть достигнуты путем идентификации ключевых регуляторных генов на больших фенотипических скринингах генетически разнообразных картирующих популяций, полученных естественным или экспериментальным путем.
Культуры можно условно разделить на два класса: культуры, выращиваемые для получения общей биомассы, и культуры, выращиваемые для производства семян или фруктов. Что касается последнего, влияние абиотических и биотических стрессов на урожай следует исследовать как на вегетативной, так и на генеративной фазах жизненного цикла растения, поскольку оба они сильно связаны с репродуктивным успехом. Температурный стресс является одним из наиболее пагубных абиотических стрессов во время генеративной фазы, и для многих видов сообщалось о снижении урожайности из-за высоких температур во время цветения (Hedhly, 2011; Bita and Gerats, 2013). Тепловой стресс во время цветения сельскохозяйственных культур уже оказывает большое влияние на мировое производство продуктов питания, и это воздействие усилится в ближайшие годы из-за ожидаемых климатических изменений. Например, в 2000 году 15% мировых посевов кукурузы ( Zea mays ) подвергались как минимум пятидневному тепловому стрессу в период цветения, а к 2050 году ожидается увеличение этого показателя до 44% ( Gourdji et al., 2013). Помимо кукурузы, многие другие зерновые, такие как пшеница ( Triticum aestivum ), ячмень ( Hordeum vulgare ) и рис ( Oryza sativa ), подвержены воздействию высоких температур во время репродуктивной фазы (Barnabás et al., 2008). О негативных последствиях теплового стресса сообщалось также для других важных культур из различных семейств растений, таких как томат ( Solanum lycopersicum ) (Sato et al., 2002, 2006) и перец ( Capsicum annuum ) (Marcelis et al. , 2004) из семейства Solanaceae , канола и горчица ( Brassica juncea , Brassica rapa и Brassica napus ) (Angadi et al.
Тепловой стресс во время цветения сельскохозяйственных культур уже оказывает большое влияние на мировое производство продуктов питания, и это воздействие усилится в ближайшие годы из-за ожидаемых климатических изменений. Например, в 2000 году 15% мировых посевов кукурузы ( Zea mays ) подвергались как минимум пятидневному тепловому стрессу в период цветения, а к 2050 году ожидается увеличение этого показателя до 44% ( Gourdji et al., 2013). Помимо кукурузы, многие другие зерновые, такие как пшеница ( Triticum aestivum ), ячмень ( Hordeum vulgare ) и рис ( Oryza sativa ), подвержены воздействию высоких температур во время репродуктивной фазы (Barnabás et al., 2008). О негативных последствиях теплового стресса сообщалось также для других важных культур из различных семейств растений, таких как томат ( Solanum lycopersicum ) (Sato et al., 2002, 2006) и перец ( Capsicum annuum ) (Marcelis et al. , 2004) из семейства Solanaceae , канола и горчица ( Brassica juncea , Brassica rapa и Brassica napus ) (Angadi et al. , 2000; Gan et al., 2004) из Brassicaceae семейство, соя ( Glycine max ) (Djanaguiraman et al., 2013) и нута ( Cicer arietinum ) (Clarke, Siddique, 2004) из семейства Fabiaceae , а также персика ( Prunus persica ) (Hedhly et al., 2005) и вишни ( Prunus avium ) ( Hedhly et al., 2004) из семейства Rosaceae . Таким образом, тепловой стресс является важным фактором, влияющим на урожайность сельскохозяйственных культур во всем мире.
, 2000; Gan et al., 2004) из Brassicaceae семейство, соя ( Glycine max ) (Djanaguiraman et al., 2013) и нута ( Cicer arietinum ) (Clarke, Siddique, 2004) из семейства Fabiaceae , а также персика ( Prunus persica ) (Hedhly et al., 2005) и вишни ( Prunus avium ) ( Hedhly et al., 2004) из семейства Rosaceae . Таким образом, тепловой стресс является важным фактором, влияющим на урожайность сельскохозяйственных культур во всем мире.
Абиотические и биотические стрессы влияют на урожай как на вегетативной, так и на генеративной фазах жизненного цикла растения, но тепловой стресс особенно пагубен на репродуктивной стадии семенных и плодовых культур (Wahid et al., 2007). Однако чувствительность к температурному стрессу меняется во время развития цветов и семян. Выявлено два чувствительных периода: во время мейоза и вокруг оплодотворения. Многие исследования, например, на злаках, томатах и Arabidopsis thaliana , показали, что развитие мужских репродуктивных органов более чувствительно к повышенным температурам, чем развитие женских репродуктивных органов (Kim et al. , 2001; Sato et al. , 2006). Эти исследования показали, что тепловой стресс препятствует раскрытию пыльников, тем самым уменьшая выделение пыльцы.Тепловой стресс также приводит к укорачиванию пыльников из-за пониженного количества ауксина (Sakata et al., 2010), что приводит к несоответствию между созреванием рыльца и пыльников, предотвращая самооплодотворение у двуполых однодомных растений. Более того, жизнеспособность пыльцы снижается из-за нарушения мужского мейоза при тепловом стрессе (Endo et al., 2009). Сообщалось не только о нарушении развития мужских органов и пыльцы, но и об изменениях в развитии женских органов.Помимо уродства семяпочки и окружающих тканей (Hedhly, 2011), описаны изменения восприимчивости рыльца, приводящие к уменьшению количества пыльцевых трубок, достигающих яичника (Saini and Aspinall, 1982). Время и тяжесть стресса во многом определяют, какие физиологические процессы нарушаются. Однако ответы у многих видов схожи, даже если они не являются близкородственными (Hedhly, 2011).
, 2001; Sato et al. , 2006). Эти исследования показали, что тепловой стресс препятствует раскрытию пыльников, тем самым уменьшая выделение пыльцы.Тепловой стресс также приводит к укорачиванию пыльников из-за пониженного количества ауксина (Sakata et al., 2010), что приводит к несоответствию между созреванием рыльца и пыльников, предотвращая самооплодотворение у двуполых однодомных растений. Более того, жизнеспособность пыльцы снижается из-за нарушения мужского мейоза при тепловом стрессе (Endo et al., 2009). Сообщалось не только о нарушении развития мужских органов и пыльцы, но и об изменениях в развитии женских органов.Помимо уродства семяпочки и окружающих тканей (Hedhly, 2011), описаны изменения восприимчивости рыльца, приводящие к уменьшению количества пыльцевых трубок, достигающих яичника (Saini and Aspinall, 1982). Время и тяжесть стресса во многом определяют, какие физиологические процессы нарушаются. Однако ответы у многих видов схожи, даже если они не являются близкородственными (Hedhly, 2011). Это делает очень актуальным изучение этих процессов на модельном растении, таком как арабидопсис.Однако удивительно, что многие исследования этого вида сосредоточены на влиянии повышенных температур на прорастание (Baskin, Baskin, 1983; Schmuths et al., 2006; He et al., 2014; Silva-Correia et al., 2014), цветение время (Balasubramanian et al., 2006; Y. Li et al., 2014) и архитектура растений (Wahid et al., 2007; Patel and Franklin, 2009; Antoun and Ouellet, 2013), но лишь очень немногие из них обращаются к влиянию тепла на плодородие (Kim et al., 2001; (Warner and Erwin, 2005; Zinn et al., 2010)
Это делает очень актуальным изучение этих процессов на модельном растении, таком как арабидопсис.Однако удивительно, что многие исследования этого вида сосредоточены на влиянии повышенных температур на прорастание (Baskin, Baskin, 1983; Schmuths et al., 2006; He et al., 2014; Silva-Correia et al., 2014), цветение время (Balasubramanian et al., 2006; Y. Li et al., 2014) и архитектура растений (Wahid et al., 2007; Patel and Franklin, 2009; Antoun and Ouellet, 2013), но лишь очень немногие из них обращаются к влиянию тепла на плодородие (Kim et al., 2001; (Warner and Erwin, 2005; Zinn et al., 2010)
У многих видов были идентифицированы теплостойкие и чувствительные к теплу сорта.Кроме того, в исследовании Arabidopsis сообщается о естественных вариациях теплового отклика во время цветения (Warner and Erwin, 2005). К сожалению, о генетических факторах, вызывающих такое изменение теплового отклика, известно гораздо меньше. Чтобы определить механизмы, ответственные за термостойкость, термостойкие и чувствительные сорта или различные природные образцы можно сравнить на физиологическом, молекулярном и генетическом уровнях. У томатов профили экспрессии метаболитов и генов сравнивали у сортов с разным уровнем термостойкости.В пыльниках толерантных сортов наблюдалась более высокая конститутивная экспрессия белков теплового шока и факторов транскрипции теплового шока (Bita et al., 2011). Более того, сообщалось о более высокой активности инвертазы при тепловом стрессе у толерантных сортов (Dorion et al., 1996; Li et al., 2012), а у чувствительных сортов наблюдалось подавление переносчика пролина (Sato et al., 2006). Эта информация может быть полезна для понимания механизмов толерантности, но чтобы быть полезной для селекции термостойких культур, необходимо идентифицировать аллельные вариации генов, способствующих термостойкости.
У томатов профили экспрессии метаболитов и генов сравнивали у сортов с разным уровнем термостойкости.В пыльниках толерантных сортов наблюдалась более высокая конститутивная экспрессия белков теплового шока и факторов транскрипции теплового шока (Bita et al., 2011). Более того, сообщалось о более высокой активности инвертазы при тепловом стрессе у толерантных сортов (Dorion et al., 1996; Li et al., 2012), а у чувствительных сортов наблюдалось подавление переносчика пролина (Sato et al., 2006). Эта информация может быть полезна для понимания механизмов толерантности, но чтобы быть полезной для селекции термостойких культур, необходимо идентифицировать аллельные вариации генов, способствующих термостойкости.
Естественные различия в термостойкости между сортами можно использовать для обнаружения полезных аллелей при разделении картируемых популяций. Более того, такие исследования выявляют фактические генетические регуляторы, управляющие определенным ответом, в отличие от выявления транскрипционных различий при тепловом стрессе в исследованиях экспрессии генов, которые выявляют многие последующие эффекты ответа. Были разработаны популяции для двух родительского картирования, которые используются для картирования локусов количественных признаков (QTL) термостойкости различных культур, таких как рис (Ye et al., 2012), вигну ( Vigna unguiculata ) (Lucas et al., 2013) и томат (Grilli et al., 2007). Эти исследования выявили несколько QTL (от 2 до 18) с объясненной дисперсией от 2 до 20%. Эти исследования показывают, что анализ QTL и последующее точное картирование и клонирование являются многообещающим способом идентификации генов термостойкости. Кроме того, количество QTL, обнаруженных в этих исследованиях, и относительно низкая объясненная дисперсия для большинства из них указывают на то, что термостойкость во время цветения является мультигенным признаком.К сожалению, ни одно из упомянутых исследований не выявило основных причинных генов. Такая идентификация в классических исследованиях сцепления занимает много времени из-за обширных областей, связанных с интересующей характеристикой. Поэтому картирование полногеномных ассоциаций (GWA) в естественных популяциях может быть очень полезным, потому что из-за быстрого распада сцепления разрешение намного выше, и поэтому точное картирование часто не требуется для идентификации генов-кандидатов (Bergelson and Roux, 2010).
Были разработаны популяции для двух родительского картирования, которые используются для картирования локусов количественных признаков (QTL) термостойкости различных культур, таких как рис (Ye et al., 2012), вигну ( Vigna unguiculata ) (Lucas et al., 2013) и томат (Grilli et al., 2007). Эти исследования выявили несколько QTL (от 2 до 18) с объясненной дисперсией от 2 до 20%. Эти исследования показывают, что анализ QTL и последующее точное картирование и клонирование являются многообещающим способом идентификации генов термостойкости. Кроме того, количество QTL, обнаруженных в этих исследованиях, и относительно низкая объясненная дисперсия для большинства из них указывают на то, что термостойкость во время цветения является мультигенным признаком.К сожалению, ни одно из упомянутых исследований не выявило основных причинных генов. Такая идентификация в классических исследованиях сцепления занимает много времени из-за обширных областей, связанных с интересующей характеристикой. Поэтому картирование полногеномных ассоциаций (GWA) в естественных популяциях может быть очень полезным, потому что из-за быстрого распада сцепления разрешение намного выше, и поэтому точное картирование часто не требуется для идентификации генов-кандидатов (Bergelson and Roux, 2010). ). Поскольку аналогичные тепловые реакции наблюдаются у многих видов, гены термостойкости, идентифицированные GWA-картированием в естественной популяции арабидопсиса, вероятно, будут иметь гомологичные эквиваленты в сельскохозяйственных культурах.Учитывая различия в чувствительности к тепловому стрессу, наблюдаемые для разных стадий развития и генотипов у разных видов, настало время объединить анализ этих наблюдений в одном исследовании. Это дает возможность идентифицировать естественные вариации ключевых регуляторов в ответ на тепловой стресс, которые проявляют свою функцию в определенной фазе развития. Эти идеи могут обеспечить более глубокое понимание регуляции времени развития и естественных стратегий и механизмов, с помощью которых растения справляются с абиотическими стрессами.
). Поскольку аналогичные тепловые реакции наблюдаются у многих видов, гены термостойкости, идентифицированные GWA-картированием в естественной популяции арабидопсиса, вероятно, будут иметь гомологичные эквиваленты в сельскохозяйственных культурах.Учитывая различия в чувствительности к тепловому стрессу, наблюдаемые для разных стадий развития и генотипов у разных видов, настало время объединить анализ этих наблюдений в одном исследовании. Это дает возможность идентифицировать естественные вариации ключевых регуляторов в ответ на тепловой стресс, которые проявляют свою функцию в определенной фазе развития. Эти идеи могут обеспечить более глубокое понимание регуляции времени развития и естественных стратегий и механизмов, с помощью которых растения справляются с абиотическими стрессами.
Мы стремились систематически исследовать генетические основы термостойкости на различных этапах развития репродукции Arabidopsis. Не все стадии развития цветов и семян были одинаково чувствительны, и мы смогли различить тепловые реакции до и после цветения. Картирование GWA было выполнено на данных, полученных для различных областей вдоль соцветия, что привело к появлению QTL, специфичных для стадии развития. Можно было наблюдать сильные ассоциации для пред- и постантезисного теплового ответа, и было идентифицировано несколько генов-кандидатов, которые ранее не были связаны с тепловым стрессом.Некоторые из этих генов-кандидатов могут быть функционально подтверждены с помощью нокаут-анализов вставок Т-ДНК и искусственно индуцированных мутантов, что обеспечивает уверенность в применяемых методах.
Картирование GWA было выполнено на данных, полученных для различных областей вдоль соцветия, что привело к появлению QTL, специфичных для стадии развития. Можно было наблюдать сильные ассоциации для пред- и постантезисного теплового ответа, и было идентифицировано несколько генов-кандидатов, которые ранее не были связаны с тепловым стрессом.Некоторые из этих генов-кандидатов могут быть функционально подтверждены с помощью нокаут-анализов вставок Т-ДНК и искусственно индуцированных мутантов, что обеспечивает уверенность в применяемых методах.
РЕЗУЛЬТАТЫ
Для исследования естественных вариаций снижения плодородия, вызванных кратковременным повышением температуры, коллекция природных образцов Arabidopsis была подвергнута тепловому стрессу и впоследствии использована для картирования GWA.
Количественная оценка реакции на тепловой стресс путем определения длины кремнезема
Цветущие растения широкого спектра природных образцов Arabidopsis подвергали 1-дневной обработке при 35 ° C. Во-первых, мы проверили, можно ли количественно оценить влияние теплового стресса, взяв длину каждого стручка вдоль основного соцветия в качестве меры количества семян. Образец Di-1 (происхождение Дижон, Франция), который показал промежуточный отклик на термическую обработку, был выбран для подробного анализа длины стручка, размера семян и количества семян на стручок для всех стручков вдоль основного соцветия (рис. 1). Наблюдалась линейная корреляция между длиной кремнезема и количеством семян на кремнезем (Рисунок 1C; r 2 = 0.90). О такой высокой корреляции также сообщалось ранее для образцов Landsberg erecta и островов Зеленого Мыса (Alonso-Blanco et al., 1999). Уменьшение длины кремнезема в основном вызвано меньшим количеством семян на кремний, чем меньшим размером семян. Обычно наблюдались два типа семян: большинство светло-коричневых семян и меньшее количество темно-коричневых мелких семян (рис. 1E). Тесты на всхожесть показали, что большинство мелких темно-коричневых семян не прорастают, в то время как почти все светло-коричневые семена прорастают (рис.
Во-первых, мы проверили, можно ли количественно оценить влияние теплового стресса, взяв длину каждого стручка вдоль основного соцветия в качестве меры количества семян. Образец Di-1 (происхождение Дижон, Франция), который показал промежуточный отклик на термическую обработку, был выбран для подробного анализа длины стручка, размера семян и количества семян на стручок для всех стручков вдоль основного соцветия (рис. 1). Наблюдалась линейная корреляция между длиной кремнезема и количеством семян на кремнезем (Рисунок 1C; r 2 = 0.90). О такой высокой корреляции также сообщалось ранее для образцов Landsberg erecta и островов Зеленого Мыса (Alonso-Blanco et al., 1999). Уменьшение длины кремнезема в основном вызвано меньшим количеством семян на кремний, чем меньшим размером семян. Обычно наблюдались два типа семян: большинство светло-коричневых семян и меньшее количество темно-коричневых мелких семян (рис. 1E). Тесты на всхожесть показали, что большинство мелких темно-коричневых семян не прорастают, в то время как почти все светло-коричневые семена прорастают (рис.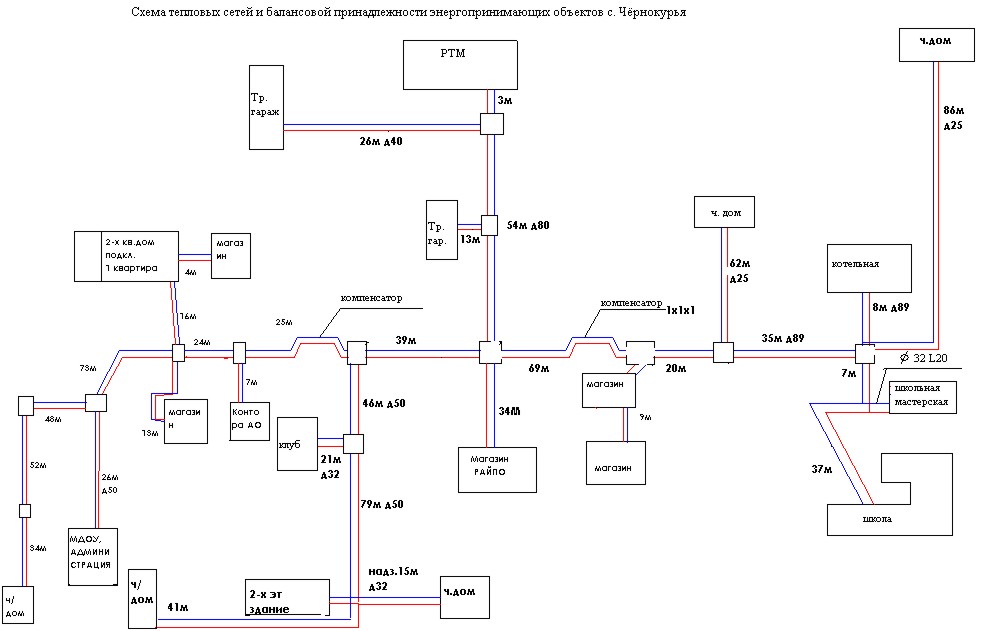 1D).Мелкие темно-коричневые семена в основном присутствовали в стручках, эмбриогенез которых уже начался в день обработки. Следовательно, эти маленькие темно-коричневые семена, скорее всего, являются абортированными семенами. Эти предполагаемые аборты семян не привели к получению более коротких стручков, что указывает на то, что длина стручков является мерой успешного оплодотворения, приводящего к инициации эмбриогенеза, и не всегда может отражать количество жизнеспособных семян в стручках. Стволовые растения контрольных растений также содержали несколько маленьких темно-коричневых семян (рис. 1А), что указывает на то, что прерывание семени не только является результатом теплового стресса, но также может быть вызвано другими факторами.
1D).Мелкие темно-коричневые семена в основном присутствовали в стручках, эмбриогенез которых уже начался в день обработки. Следовательно, эти маленькие темно-коричневые семена, скорее всего, являются абортированными семенами. Эти предполагаемые аборты семян не привели к получению более коротких стручков, что указывает на то, что длина стручков является мерой успешного оплодотворения, приводящего к инициации эмбриогенеза, и не всегда может отражать количество жизнеспособных семян в стручках. Стволовые растения контрольных растений также содержали несколько маленьких темно-коричневых семян (рис. 1А), что указывает на то, что прерывание семени не только является результатом теплового стресса, но также может быть вызвано другими факторами.
стручка, количество семян и размер семян на стручок вдоль основного соцветия растений Di-1 и скорость прорастания семян.
(A) и (B) Сравнение длины стручка и количества семян (светло- и темно-коричневых) в контроле ( n = 3 для размера семян; n = 5 для длины кремния) и тепла -обработанные ( n = 5) условиях ( [A] и [B] , соответственно).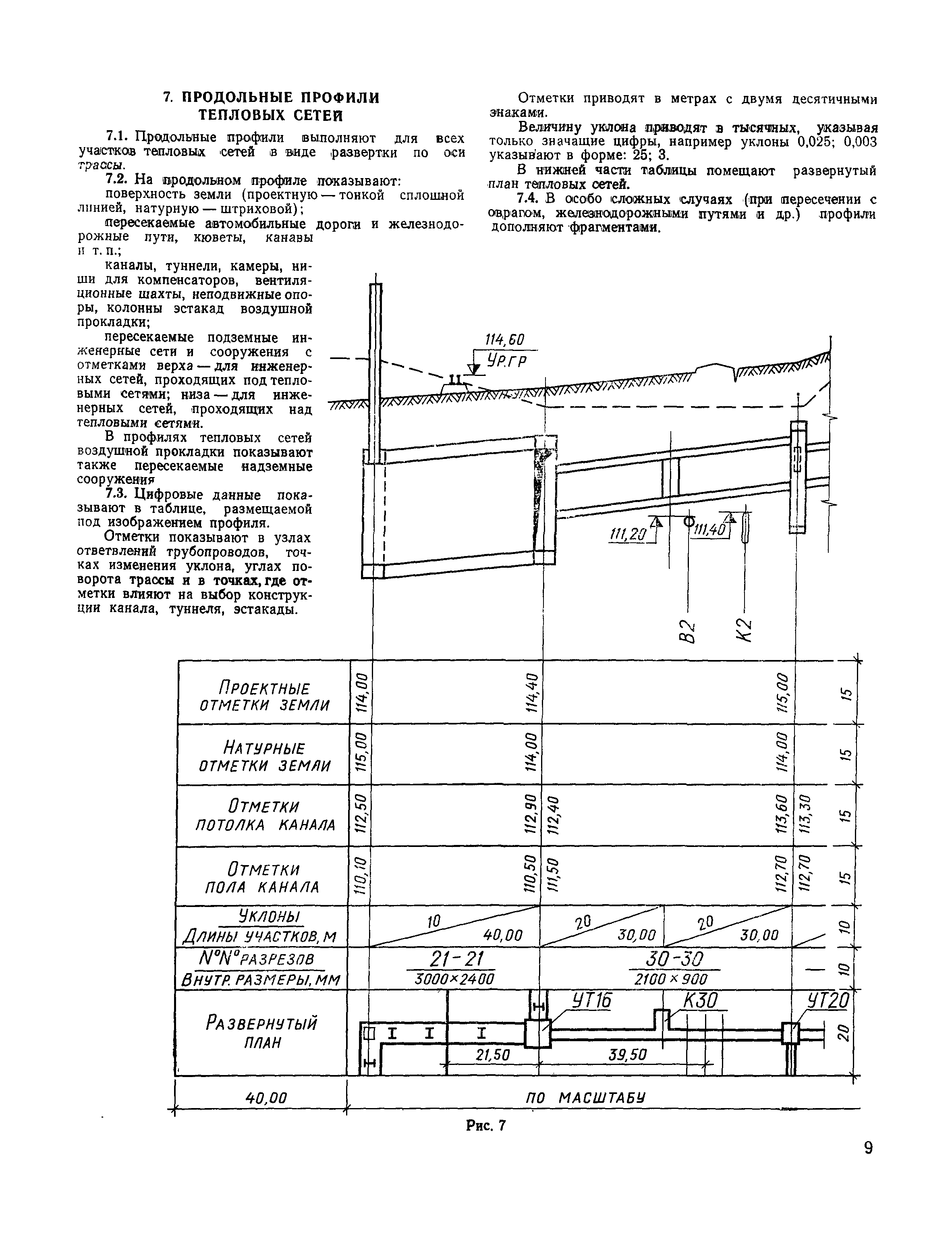 Планки погрешностей представляют SD.Номера позиций относятся к цветку, который открылся первым в день обработки, который получил номер позиции 0.
Планки погрешностей представляют SD.Номера позиций относятся к цветку, который открылся первым в день обработки, который получил номер позиции 0.
(C) Линейная корреляция между длиной стручка и количеством семян на стручке на основе данных, представленных в (A). и (В) .
(D) Всхожесть светло-коричневых ( n = 213) и темно-коричневых ( n = 244) семян термически обработанных растений.
(E) Гранулометрический состав светло-коричневых ( n = 575) и темно-коричневых ( n = 526) семян термически обработанных растений.
Цветок, который открылся первым в день обработки, был помечен для каждой повторности из 285 образцов, использованных в эксперименте. Этот тег позволил нам связать эффект теплового стресса со стадией развития цветков и стручков вдоль соцветия (рис. 2). Что касается метки, все стручки получили номер относительного положения, и впоследствии мы определили пять областей развития, от A до E. Чтобы связать наблюдаемое уменьшение длины стручка с нарушением развития цветков и эмбриогенеза, мы сопоставили регионы, как показано на рисунке 2 с данными о времени и порядке событий, происходящих во время этих процессов развития у Arabidopsis (Таблица 1; Smyth et al., 1990; Мэнсфилд и Бриарти, 1991; Schneitz et al., 1995; Sanders et al., 1999). Область A (номера позиций кремния от -20 до -10) связана с поздним эмбриогенезом. Область B (номера позиций кремния от -10 до 0) связана с процессами, происходящими вскоре после цветения, такими как оплодотворение и ранний эмбриогенез. Область C (номера кремниевых позиций от 0 до 10) связана с процессами непосредственно перед цветением, такими как удлинение пыльника и женский мейоз, а область D (кремнистые позиции с номерами от 10 до 20) связана с процессами, которые происходят за несколько дней до цветения, как и у самцов. мейоз.И, наконец, область E (номера позиций силикона от 20 до 30) соответствует раннему развитию цветка. Хотя мы определили каждый регион по фиксированному количеству стручков, не делая различия между образцами, мы отмечаем, что наблюдались естественные вариации количества цветков, раскрывающихся за день.
Чтобы связать наблюдаемое уменьшение длины стручка с нарушением развития цветков и эмбриогенеза, мы сопоставили регионы, как показано на рисунке 2 с данными о времени и порядке событий, происходящих во время этих процессов развития у Arabidopsis (Таблица 1; Smyth et al., 1990; Мэнсфилд и Бриарти, 1991; Schneitz et al., 1995; Sanders et al., 1999). Область A (номера позиций кремния от -20 до -10) связана с поздним эмбриогенезом. Область B (номера позиций кремния от -10 до 0) связана с процессами, происходящими вскоре после цветения, такими как оплодотворение и ранний эмбриогенез. Область C (номера кремниевых позиций от 0 до 10) связана с процессами непосредственно перед цветением, такими как удлинение пыльника и женский мейоз, а область D (кремнистые позиции с номерами от 10 до 20) связана с процессами, которые происходят за несколько дней до цветения, как и у самцов. мейоз.И, наконец, область E (номера позиций силикона от 20 до 30) соответствует раннему развитию цветка. Хотя мы определили каждый регион по фиксированному количеству стручков, не делая различия между образцами, мы отмечаем, что наблюдались естественные вариации количества цветков, раскрывающихся за день. Несмотря на то, что такие фазовые сдвиги в сроках развития могут вносить шум, это не повлияло отрицательно на последующий анализ, вероятно, потому, что размер назначенных регионов относительно велик по сравнению с наблюдаемыми различиями между образцами.Таким образом, мы предполагаем, что процессы развития, относящиеся к различным регионам, верны для большинства образцов и анализов, описанных ниже.
Несмотря на то, что такие фазовые сдвиги в сроках развития могут вносить шум, это не повлияло отрицательно на последующий анализ, вероятно, потому, что размер назначенных регионов относительно велик по сравнению с наблюдаемыми различиями между образцами.Таким образом, мы предполагаем, что процессы развития, относящиеся к различным регионам, верны для большинства образцов и анализов, описанных ниже.
Очертание главного соцветия.
Схема сфотографированных соцветий репрезентативного восприимчивого образца, выращенного в условиях контроля (A) и термообработанных (B) . Определение номера позиции и регионов развития; кремний с меткой произошел от цветка, который распустился первым в день обработки.Номера позиций указаны относительно тега. Были определены пять регионов (от A до E), каждая из которых содержит десять стеблей и представляет различные стадии развития, как показано в таблице 1.
Таблица 1. Сопоставление знаковых событий в развитии цветов и семян с областями развития вдоль соцветия, как показано на рисунке 2Естественная изменчивость теплового ответа зависит от стадии развития
Естественная изменчивость завязки семян при тепловом стрессе регистрировалась путем измерения длины всех стручков вдоль основного соцветия каждого отдельного растения всех образцов.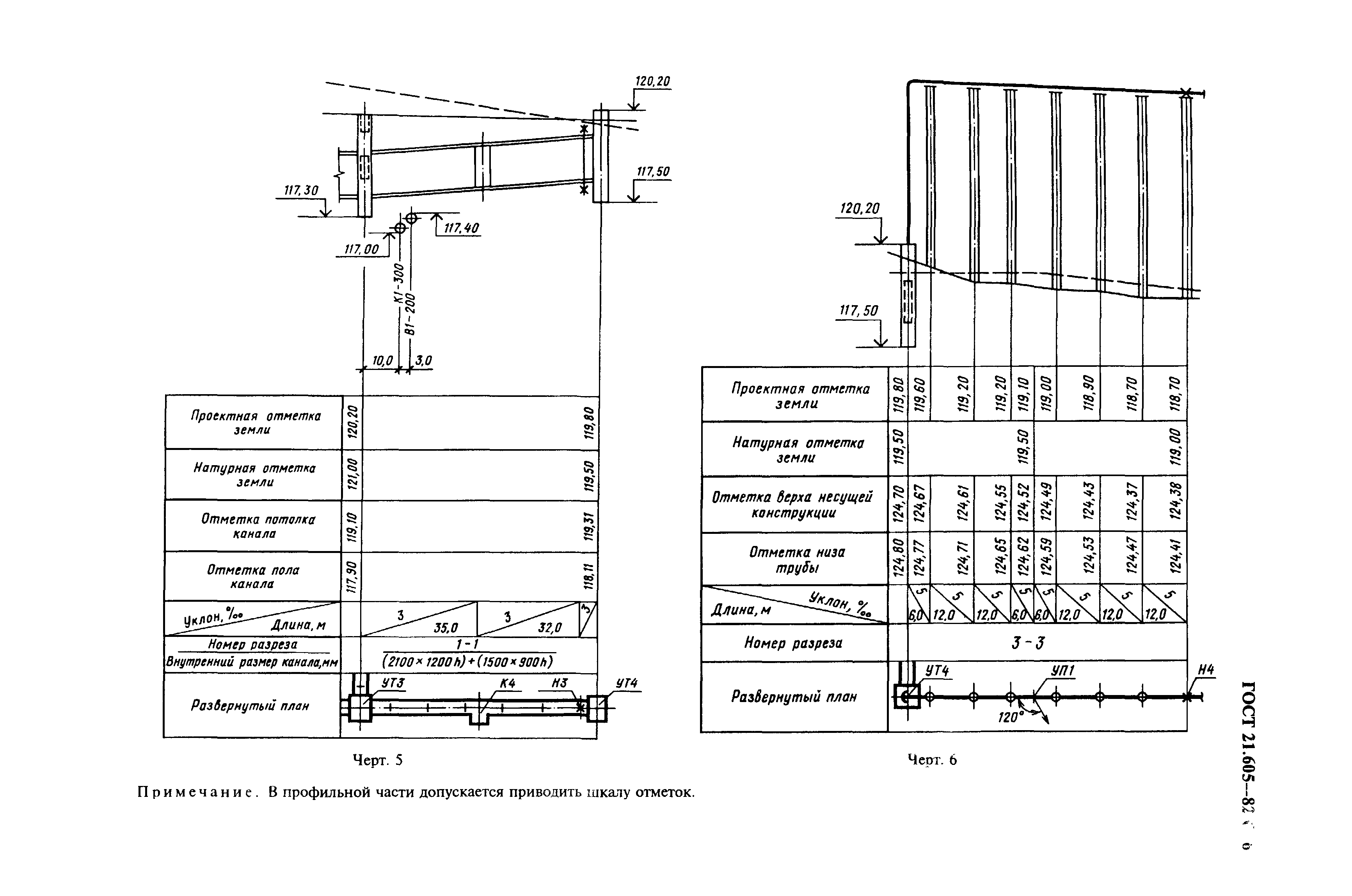 Чтобы определить общую картину ответа, было рассчитано среднее значение длины кремнистой железы всех растений в эксперименте для всех позиций кремния вдоль основного соцветия (рис. 3А). Эффект от обработки варьировался по соцветию. Например, область A (поздний эмбриогенез) и область E (раннее развитие цветков) меньше всего пострадали от тепловой обработки у всех образцов (рис. 3A), в то время как стручки в других регионах были, по крайней мере, затронуты у некоторых образцов. После термообработки наблюдались два минимума общей средней длины кремнезема.Первый минимум был обнаружен для кремнеземов в фазе раннего эмбриогенеза (область B) с абсолютным минимумом в позиции минус три. В среднем длина стручка в этом месте уменьшилась с 13,4 до 8,2 мм, что соответствует уменьшению в среднем примерно на 21 семя на стручок (рис. 1С). Второй минимум охватывал большую область, в которой происходит мужской и женский мейоз. Наименьшее значение было достигнуто в позиции десять со средним уменьшением длины кремнезема с 13,0 до 6,3 мм, что соответствует уменьшению в среднем примерно на 27 семян на стручок (Рисунок 1C).
Чтобы определить общую картину ответа, было рассчитано среднее значение длины кремнистой железы всех растений в эксперименте для всех позиций кремния вдоль основного соцветия (рис. 3А). Эффект от обработки варьировался по соцветию. Например, область A (поздний эмбриогенез) и область E (раннее развитие цветков) меньше всего пострадали от тепловой обработки у всех образцов (рис. 3A), в то время как стручки в других регионах были, по крайней мере, затронуты у некоторых образцов. После термообработки наблюдались два минимума общей средней длины кремнезема.Первый минимум был обнаружен для кремнеземов в фазе раннего эмбриогенеза (область B) с абсолютным минимумом в позиции минус три. В среднем длина стручка в этом месте уменьшилась с 13,4 до 8,2 мм, что соответствует уменьшению в среднем примерно на 21 семя на стручок (рис. 1С). Второй минимум охватывал большую область, в которой происходит мужской и женский мейоз. Наименьшее значение было достигнуто в позиции десять со средним уменьшением длины кремнезема с 13,0 до 6,3 мм, что соответствует уменьшению в среднем примерно на 27 семян на стручок (Рисунок 1C). В целом наблюдались большие различия в ответе на обработку, начиная от образцов без четкого ответа до полного отсутствия силикатов; следовательно, два минимума не всегда обнаруживались во всех образцах. Например, образец C24 (происхождение из Португалии) не показал уменьшения длины силикона ни в одном месте, тогда как образец Rmx-A180 (происхождение из Мичигана) явно пострадал в обоих регионах. Кроме того, образец Arby-1 (происхождение из Швеции) сильно пострадал по длине кремнезема вдоль большей части соцветия (Рисунки 3B — 3D).Различия в длине кремнезема в контрольных условиях и в условиях теплового стресса были высокими между образцами, тогда как колебания внутри образцов были низкими. Это привело к высокой наследуемости в широком смысле (H 2 от 0,58 до 0,73; Таблица 2), что указывает на то, что наблюдаемые различия в длине стручка между образцами по большей части определяются генетическими факторами. Кроме того, между разными образцами наблюдались существенные различия в общем количестве цветков, сформировавшихся вдоль основного соцветия в репродуктивный период (рис.
В целом наблюдались большие различия в ответе на обработку, начиная от образцов без четкого ответа до полного отсутствия силикатов; следовательно, два минимума не всегда обнаруживались во всех образцах. Например, образец C24 (происхождение из Португалии) не показал уменьшения длины силикона ни в одном месте, тогда как образец Rmx-A180 (происхождение из Мичигана) явно пострадал в обоих регионах. Кроме того, образец Arby-1 (происхождение из Швеции) сильно пострадал по длине кремнезема вдоль большей части соцветия (Рисунки 3B — 3D).Различия в длине кремнезема в контрольных условиях и в условиях теплового стресса были высокими между образцами, тогда как колебания внутри образцов были низкими. Это привело к высокой наследуемости в широком смысле (H 2 от 0,58 до 0,73; Таблица 2), что указывает на то, что наблюдаемые различия в длине стручка между образцами по большей части определяются генетическими факторами. Кроме того, между разными образцами наблюдались существенные различия в общем количестве цветков, сформировавшихся вдоль основного соцветия в репродуктивный период (рис.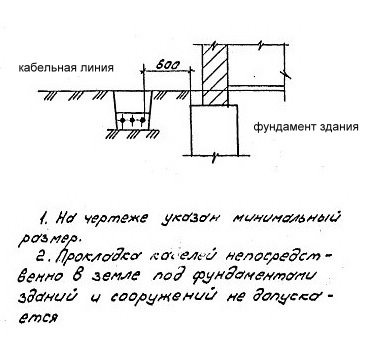 4А).У большинства образцов количество цветков, образовавшихся на термообработанных растениях, было выше, чем в контрольных условиях (рис. 4В). Среднее количество цветков, образовавшихся на всех термообработанных растениях, также было значительно больше, чем среднее количество цветков в контрольных растениях (P <0,001). В среднем у термически обработанных растений появилось на 12% больше цветков, чем у растений в контрольных условиях, что указывает на механизм компенсации потери жизнеспособных семян.
4А).У большинства образцов количество цветков, образовавшихся на термообработанных растениях, было выше, чем в контрольных условиях (рис. 4В). Среднее количество цветков, образовавшихся на всех термообработанных растениях, также было значительно больше, чем среднее количество цветков в контрольных растениях (P <0,001). В среднем у термически обработанных растений появилось на 12% больше цветков, чем у растений в контрольных условиях, что указывает на механизм компенсации потери жизнеспособных семян.
Silique вдоль основных соцветий контрольных (синие) и термически обработанных (красный) растений (Среднее ± стандартное отклонение).
(A) Общее среднее значение для всех растений в эксперименте.
(B) Репрезентативный толерантный образец, C24.
(C) Представитель умеренно чувствительного образца Rmx-A180.
(D) Представитель чрезвычайно чувствительного образца Arby-1.
(B) от до (D) Контроль, n = 3; обработка, n = 5. Столбцы указывают sd длины кремния среди повторов, а сплошные линии указывают средние значения.
Таблица 2. Наследуемость длины стручка в широком смысле в условиях контроля и теплового стресса для каждой области развития (B, C и D, как определено на рисунке 2) и времени цветения Рисунок 4.Сравнение общего количества и соотношения цветков вдоль Основное соцветие у контрольных и термообработанных растений у всех образцов.
(A) Частотное распределение общего числа цветков на главное соцветие, усредненное на образец.
(B) Частотное распределение отношения количества цветков вдоль основного соцветия в термообработанных и контрольных условиях на образец.
Для количественной оценки воздействия тепла на различные стадии развития была рассчитана средняя длина кремнезема на область развития для контрольных и термообработанных растений. Области A и E не учитывались, поскольку у некоторых образцов с коротким основным соцветием не все позиции в этих регионах развили цветок. Кроме того, мы наблюдали, что после определения окна чувствительности развития для большинства образцов эти области находились за пределами этого окна. Для количественной оценки воздействия тепла независимо от размера кремнезема в контрольных условиях простые остатки длины кремнезема контрольных и термообработанных растений были рассчитаны с использованием линейной регрессии.Для всех этих признаков фертильности был проведен корреляционный анализ, чтобы определить, связаны ли они между собой (таблица 3). Время цветения также было включено в этот корреляционный анализ. Значительные положительные корреляции были обнаружены между длиной стеблей термообработанных растений в регионах B, C и D. Это указывает на то, что растения, чувствительные в области B в целом, также чувствительны в областях C и D и наоборот. Хотя тепловые реакции в разных регионах могут быть взаимозависимыми, корреляция не всегда отражает прямую причинную связь и может быть результатом последующих эффектов ранних событий.
Области A и E не учитывались, поскольку у некоторых образцов с коротким основным соцветием не все позиции в этих регионах развили цветок. Кроме того, мы наблюдали, что после определения окна чувствительности развития для большинства образцов эти области находились за пределами этого окна. Для количественной оценки воздействия тепла независимо от размера кремнезема в контрольных условиях простые остатки длины кремнезема контрольных и термообработанных растений были рассчитаны с использованием линейной регрессии.Для всех этих признаков фертильности был проведен корреляционный анализ, чтобы определить, связаны ли они между собой (таблица 3). Время цветения также было включено в этот корреляционный анализ. Значительные положительные корреляции были обнаружены между длиной стеблей термообработанных растений в регионах B, C и D. Это указывает на то, что растения, чувствительные в области B в целом, также чувствительны в областях C и D и наоборот. Хотя тепловые реакции в разных регионах могут быть взаимозависимыми, корреляция не всегда отражает прямую причинную связь и может быть результатом последующих эффектов ранних событий. Кроме того, наблюдалась сильная отрицательная корреляция между временем цветения и длиной стерни у термически обработанных растений (Таблица 3). Для синхронизации цветения растения яровировали в течение 10 недель, но разница во времени цветения все еще составляла 4 недели между самым ранним и последним цветущими образцами (дополнительный набор данных 1). Следовательно, термообработку проводили партиями, чтобы сохранить одинаковый временной интервал между началом цветения и обработкой как для ранне, так и для позднецветущих образцов.Корреляция между временем цветения и тепловой реакцией также предполагает роль времени развития в чувствительности к тепловому стрессу. Что касается теплового отклика, мы также наблюдали высокую наследуемость по времени цветения (H 2 = 0,96; Таблица 2).
Кроме того, наблюдалась сильная отрицательная корреляция между временем цветения и длиной стерни у термически обработанных растений (Таблица 3). Для синхронизации цветения растения яровировали в течение 10 недель, но разница во времени цветения все еще составляла 4 недели между самым ранним и последним цветущими образцами (дополнительный набор данных 1). Следовательно, термообработку проводили партиями, чтобы сохранить одинаковый временной интервал между началом цветения и обработкой как для ранне, так и для позднецветущих образцов.Корреляция между временем цветения и тепловой реакцией также предполагает роль времени развития в чувствительности к тепловому стрессу. Что касается теплового отклика, мы также наблюдали высокую наследуемость по времени цветения (H 2 = 0,96; Таблица 2).
QTL, связанных с Специфические для стадии тепловые реакции
Для поиска генетической вариации, которая вызывает фенотипические вариации, наблюдаемые в признаках фертильности, мы выполнили GWA-картирование на данных контрольных и термообработанных растений, а также для остатков. Анализируемые признаки включали среднюю длину стручка в регионах B, C и D, а также время цветения. Картирование GWA обнаружило 15 прочно связанных однонуклеотидных полиморфизмов (SNP) с –log (значение P) выше произвольно установленного порога значимости 5,5 (Рисунок 5). Кроме того, было идентифицировано 297 умеренно ассоциированных SNP с –log (значение P) от 4 до 5,5 (дополнительная таблица 2). Произвольный порог –log (значение P) = 5.5 был установлен, чтобы сосредоточиться на SNP с наибольшей объясненной дисперсией и минимизировать вероятность обнаружения ложных срабатываний.Некоторые из этих значимых SNP находятся в неравновесном сцеплении друг с другом и, следовательно, с большой вероятностью представляют один локус. Это привело к семи уникальным основным QTL: три, относящиеся к длине кремния в контрольных условиях, и четыре, связанные с тепловым откликом (Таблица 4). Изменение длины кремнезема в результате теплового стресса частично можно объяснить QTL2, QTL4, QTL5 и QTL6. Общая объясненная фенотипическая дисперсия этих четырех QTL достигла 35,7%, что является относительно высоким показателем для исследований GWA, особенно с учетом того, что это может быть заниженной оценкой из-за возможного присутствия нескольких аллелей на локус.
Анализируемые признаки включали среднюю длину стручка в регионах B, C и D, а также время цветения. Картирование GWA обнаружило 15 прочно связанных однонуклеотидных полиморфизмов (SNP) с –log (значение P) выше произвольно установленного порога значимости 5,5 (Рисунок 5). Кроме того, было идентифицировано 297 умеренно ассоциированных SNP с –log (значение P) от 4 до 5,5 (дополнительная таблица 2). Произвольный порог –log (значение P) = 5.5 был установлен, чтобы сосредоточиться на SNP с наибольшей объясненной дисперсией и минимизировать вероятность обнаружения ложных срабатываний.Некоторые из этих значимых SNP находятся в неравновесном сцеплении друг с другом и, следовательно, с большой вероятностью представляют один локус. Это привело к семи уникальным основным QTL: три, относящиеся к длине кремния в контрольных условиях, и четыре, связанные с тепловым откликом (Таблица 4). Изменение длины кремнезема в результате теплового стресса частично можно объяснить QTL2, QTL4, QTL5 и QTL6. Общая объясненная фенотипическая дисперсия этих четырех QTL достигла 35,7%, что является относительно высоким показателем для исследований GWA, особенно с учетом того, что это может быть заниженной оценкой из-за возможного присутствия нескольких аллелей на локус. Однако большая доля отсутствующей наследственности указывает на то, что другие генетические факторы, возможно, с большими эффектами, остались необнаруженными, например, из-за низких частот аллелей или чрезмерной коррекции структуры популяции. Ни один из значимых SNP не был идентифицирован в данных из контрольных условий, что позволяет предположить, что наблюдаемое изменение длины кремнезема, объясняемое этими SNP, связано исключительно с термической обработкой. QTL с самой сильной ассоциацией и наибольшей величиной эффекта (QTL2) состоит из четырех значимых SNP, расположенных на вершине хромосомы 2 в области 0.7 кб (таблица 4). Все четыре SNP были прочно связаны с областями C и D, но не с областью B. Это указывает на то, что этот QTL является специфическим для наблюдаемой вариации средней длины кремния в положении второго минимума и, следовательно, может быть связан с процессами предантезиса, такими как как мужской, так и женский мейоз (таблица 1). QTL5 и QTL6 были специфичными для области развития C.
Однако большая доля отсутствующей наследственности указывает на то, что другие генетические факторы, возможно, с большими эффектами, остались необнаруженными, например, из-за низких частот аллелей или чрезмерной коррекции структуры популяции. Ни один из значимых SNP не был идентифицирован в данных из контрольных условий, что позволяет предположить, что наблюдаемое изменение длины кремнезема, объясняемое этими SNP, связано исключительно с термической обработкой. QTL с самой сильной ассоциацией и наибольшей величиной эффекта (QTL2) состоит из четырех значимых SNP, расположенных на вершине хромосомы 2 в области 0.7 кб (таблица 4). Все четыре SNP были прочно связаны с областями C и D, но не с областью B. Это указывает на то, что этот QTL является специфическим для наблюдаемой вариации средней длины кремния в положении второго минимума и, следовательно, может быть связан с процессами предантезиса, такими как как мужской, так и женский мейоз (таблица 1). QTL5 и QTL6 были специфичными для области развития C. Помимо женского мейоза, удлинение пыльников также имеет место на этой стадии развития во время тепловой обработки.QTL4 — единственный QTL, который, как было обнаружено, связан с первым минимумом средней длины кремния и, следовательно, связан с процессами постантеза, такими как ранний эмбриогенез. Хотя основная цель этого исследования состояла в том, чтобы идентифицировать гены, участвующие в термостойкости, собранные данные также дают представление о регуляции развития силиката в контрольных условиях. Различия, наблюдаемые между образцами в длине кремния в контрольных условиях, можно частично объяснить тремя QTL: QTL1, QTL3 и QTL7.Хотя каждый контрольный QTL был идентифицирован в отдельной области развития, умеренные ассоциации были идентифицированы по крайней мере для одной др. Области, указывая на то, что гены, которые определяют длину силикона в одной области развития, также могут играть роль в др. Регионе (Таблица 4). Для QTL1 слабая ассоциация была также обнаружена для вариации длины кремния в области B термически обработанных растений в дополнение к сильной связи в контрольных условиях, что позволяет предположить, что причинный ген влияет на длину кремнезема независимо от условий окружающей среды.
Помимо женского мейоза, удлинение пыльников также имеет место на этой стадии развития во время тепловой обработки.QTL4 — единственный QTL, который, как было обнаружено, связан с первым минимумом средней длины кремния и, следовательно, связан с процессами постантеза, такими как ранний эмбриогенез. Хотя основная цель этого исследования состояла в том, чтобы идентифицировать гены, участвующие в термостойкости, собранные данные также дают представление о регуляции развития силиката в контрольных условиях. Различия, наблюдаемые между образцами в длине кремния в контрольных условиях, можно частично объяснить тремя QTL: QTL1, QTL3 и QTL7.Хотя каждый контрольный QTL был идентифицирован в отдельной области развития, умеренные ассоциации были идентифицированы по крайней мере для одной др. Области, указывая на то, что гены, которые определяют длину силикона в одной области развития, также могут играть роль в др. Регионе (Таблица 4). Для QTL1 слабая ассоциация была также обнаружена для вариации длины кремния в области B термически обработанных растений в дополнение к сильной связи в контрольных условиях, что позволяет предположить, что причинный ген влияет на длину кремнезема независимо от условий окружающей среды. Хотя наблюдалась сильная отрицательная корреляция между временем цветения и длиной стерни у термически обработанных растений, не было обнаружено значимых ассоциаций (-log (значение P)> 5,5) для времени цветения. Тем не менее, время цветения было умеренно связано с QTL6 (-log (значение P) = 4,96). Этот QTL соответствует положению хорошо известного гена времени цветения FLOWERING LOCUS C ( FLC ), что может объяснить наблюдаемые вариации времени цветения и предполагает роль регуляции времени цветения в реакции на тепловой стресс.Для завершения нашего анализа были также собраны данные о длине кремнезема, усредненной по трем областям (B, C и D вместе), и для общего числа кремнеземов размером менее 5 мм, которые, вероятно, не содержат жизнеспособных семян. Эти признаки не зависят от возможной предвзятости, вносимой сдвигами фаз развития между образцами, и поэтому могут выявить дополнительные регуляторные локусы. Однако картирование GWA не выявило значимых ассоциаций, кроме описанных выше (дополнительный набор данных 2), что еще больше укрепило наше предположение о том, что процессы развития, относящиеся к разным регионам, верны для большинства образцов и что возможные отклонения не сильно влияют на наш анализ.
Хотя наблюдалась сильная отрицательная корреляция между временем цветения и длиной стерни у термически обработанных растений, не было обнаружено значимых ассоциаций (-log (значение P)> 5,5) для времени цветения. Тем не менее, время цветения было умеренно связано с QTL6 (-log (значение P) = 4,96). Этот QTL соответствует положению хорошо известного гена времени цветения FLOWERING LOCUS C ( FLC ), что может объяснить наблюдаемые вариации времени цветения и предполагает роль регуляции времени цветения в реакции на тепловой стресс.Для завершения нашего анализа были также собраны данные о длине кремнезема, усредненной по трем областям (B, C и D вместе), и для общего числа кремнеземов размером менее 5 мм, которые, вероятно, не содержат жизнеспособных семян. Эти признаки не зависят от возможной предвзятости, вносимой сдвигами фаз развития между образцами, и поэтому могут выявить дополнительные регуляторные локусы. Однако картирование GWA не выявило значимых ассоциаций, кроме описанных выше (дополнительный набор данных 2), что еще больше укрепило наше предположение о том, что процессы развития, относящиеся к разным регионам, верны для большинства образцов и что возможные отклонения не сильно влияют на наш анализ. .
.
Манхэттена, представляющие связи между маркерами SNP и длиной кремнезема в областях B, C и D.
(A) GWA-отображение длины кремнезема в контрольных растениях.
(B) Картирование GWA длины кремнезема в установках с термообработкой.
(C) GWA отображение остатков, представляющее тепловой отклик, не зависящий от длины кремния в контрольных условиях.
Пунктирная линия представляет порог значимости –log (значение P) = 5.5.
Таблица 4. Основные QTL, идентифицированные картированием GWA для времени цветения и средней длины кремния в областях развития B, C и D (как определено на рисунке 2) в контрольных и термически обработанных условиях и их соответствующие остатки для этих регионовПрисвоение генов-кандидатов на основе неравновесия сцепления, аннотации генов и экспрессии
Данные ресеквенирования (1001genomes.org) 502 образцов использовали для определения возможных причинных кандидатов для каждого из семи наблюдаемых QTL. Для 124 из этих 502 образцов тепловая реакция была исследована в этом исследовании. Гены в неравновесном сцеплении (LD) со значимыми SNP (порог LD> 0,4) рассматривались как кандидаты (дополнительный рисунок 1). Кроме того, аннотации генов и паттерны экспрессии генов в различных растительных тканях были получены из TAIR и браузера Arabidopsis eFP (Winter et al., 2007), соответственно. Для ряда генов-кандидатов, выбранных по вероятности и достоверности обнаружения, тепловой ответ нокаут-мутантов исследовали в двух независимых экспериментах (дополнительный набор данных 3).
Для 124 из этих 502 образцов тепловая реакция была исследована в этом исследовании. Гены в неравновесном сцеплении (LD) со значимыми SNP (порог LD> 0,4) рассматривались как кандидаты (дополнительный рисунок 1). Кроме того, аннотации генов и паттерны экспрессии генов в различных растительных тканях были получены из TAIR и браузера Arabidopsis eFP (Winter et al., 2007), соответственно. Для ряда генов-кандидатов, выбранных по вероятности и достоверности обнаружения, тепловой ответ нокаут-мутантов исследовали в двух независимых экспериментах (дополнительный набор данных 3).
Чувствительность преантезиса к тепловому стрессу
Четыре прочно связанных SNP QTL2, объясняющие вариацию длины кремния при термообработке перед протезом, связаны с девятью другими SNP, как указано в данных повторного секвенирования. Все 13 SNP расположены в интронах локуса AT2G03505 , который, следовательно, является единственным геном-кандидатом в LD со значительными SNP. AT2G03505 кодирует неизвестный белок суперсемейства углеводсвязывающих доменов Х8. Этот ген относительно высоко экспрессируется в цветках и особенно плодолистиках (Laubinger et al., 2008). В начале процесса валидации нокаутная линия Gabi-Kat была испытана на тепловую реакцию в двух независимых экспериментах. Никаких различий между тепловым откликом этой линии и Col-0 дикого типа не наблюдалось (дополнительный набор данных 3). Это говорит о том, что этот ген может не быть причинным геном, хотя сам по себе аллель Col-0 дикого типа может быть слабым или нефункциональным, и необходимы дополнительные эксперименты по нокауту и сверхэкспрессии линий не-Col-0 аллелей этого гена для окончательное определение.Поскольку причинные гены иногда также обнаруживаются за пределами интервала LD значительно ассоциированных SNP (Chao et al., 2014), среди прочего, из-за низкой связи между причинными и ассоциированными SNP (Freedman et al., 2011), другие гены в Район размером 20 т.п.н. вокруг значимых SNP (10 т.п.н. вверху и 10 т.п.н. вниз по течению) рассматривался как кандидат в соответствии со средним расстоянием сцепления в Arabidopsis, равным 10 т.
Этот ген относительно высоко экспрессируется в цветках и особенно плодолистиках (Laubinger et al., 2008). В начале процесса валидации нокаутная линия Gabi-Kat была испытана на тепловую реакцию в двух независимых экспериментах. Никаких различий между тепловым откликом этой линии и Col-0 дикого типа не наблюдалось (дополнительный набор данных 3). Это говорит о том, что этот ген может не быть причинным геном, хотя сам по себе аллель Col-0 дикого типа может быть слабым или нефункциональным, и необходимы дополнительные эксперименты по нокауту и сверхэкспрессии линий не-Col-0 аллелей этого гена для окончательное определение.Поскольку причинные гены иногда также обнаруживаются за пределами интервала LD значительно ассоциированных SNP (Chao et al., 2014), среди прочего, из-за низкой связи между причинными и ассоциированными SNP (Freedman et al., 2011), другие гены в Район размером 20 т.п.н. вокруг значимых SNP (10 т.п.н. вверху и 10 т.п.н. вниз по течению) рассматривался как кандидат в соответствии со средним расстоянием сцепления в Arabidopsis, равным 10 т. п.н. (Kim et al., 2007). Это привело к идентификации гена-кандидата, QUASIMODO-LIKE2 ( QUL2 ; AT2G03480 ), который высоко экспрессируется во время развития мужского гаметофита (Honys and Twell, 2004).Этот процесс происходит в области D, в которой наблюдалась самая высокая ассоциация. Ранее было обнаружено, что тройной мутант QUL2 и два его паралога, QUA2 ( AT1G78240 ) и QUL1 ( AT1G13860 ), имеют температурно-зависимые изменения диаметра стебля и лигнификации стебля (Fuentes et al. , 2010). Поэтому был исследован тепловой отклик линий вставки Т-ДНК QUL2 , его ближайшего паралога QUL1 и соответствующего двойного мутанта (рис. 6; дополнительный набор данных 3).Анализ экспрессии подтвердил, что одиночные и двойные мутанты QUL1 и QUL2 были истинными нокдаунами (дополнительная фигура 2). Двойной мутант был более чувствителен к нагреванию в областях C, D и E (фигура 6), но не в области B, тогда как оба одиночных мутанта напоминали дикий тип (дополнительный набор данных 3).
п.н. (Kim et al., 2007). Это привело к идентификации гена-кандидата, QUASIMODO-LIKE2 ( QUL2 ; AT2G03480 ), который высоко экспрессируется во время развития мужского гаметофита (Honys and Twell, 2004).Этот процесс происходит в области D, в которой наблюдалась самая высокая ассоциация. Ранее было обнаружено, что тройной мутант QUL2 и два его паралога, QUA2 ( AT1G78240 ) и QUL1 ( AT1G13860 ), имеют температурно-зависимые изменения диаметра стебля и лигнификации стебля (Fuentes et al. , 2010). Поэтому был исследован тепловой отклик линий вставки Т-ДНК QUL2 , его ближайшего паралога QUL1 и соответствующего двойного мутанта (рис. 6; дополнительный набор данных 3).Анализ экспрессии подтвердил, что одиночные и двойные мутанты QUL1 и QUL2 были истинными нокдаунами (дополнительная фигура 2). Двойной мутант был более чувствителен к нагреванию в областях C, D и E (фигура 6), но не в области B, тогда как оба одиночных мутанта напоминали дикий тип (дополнительный набор данных 3). Это указывает на то, что QUL1 и QUL2 , вероятно, имеют очень похожие дополнительные функции с прямым или косвенным положительным влиянием на образование кремния при нагревании.Этот результат также подчеркивает, что разные гены могут быть ответственны за тепловую реакцию до и после цветения.
Это указывает на то, что QUL1 и QUL2 , вероятно, имеют очень похожие дополнительные функции с прямым или косвенным положительным влиянием на образование кремния при нагревании.Этот результат также подчеркивает, что разные гены могут быть ответственны за тепловую реакцию до и после цветения.
Анализ мутантов.
Разница между длиной кремнезема у растений, подвергшихся тепловому стрессу, и контрольных растений (средняя длина кремния у растений теплового стресса минус средняя длина кремния у контрольных растений) дикого типа Col-0 и мутантных линий с одинарным и двойным нокдауном QUL1 и QUL2 (A) , и изогенных линий с сильным аллелем FRI и нулевым аллелем FLC (раннее цветение, FRI-flc3 ) или функциональным аллелем FLC (позднее цветение, FRI-sf2 ) (B ) .Показаны результаты одного эксперимента. Результаты второго независимого эксперимента можно найти в Supplemental Data Set 3.
Вторая значимая ассоциация (QTL5), относящаяся к стадии preanthesis, расположена в хромосомной области с очень быстрым распадом сцепления (Supplemental Figure 1). Значимый SNP не находится в LD с SNP повторной последовательности, которые расположены в восходящем направлении, а только с ограниченным количеством SNP, расположенных в 4 kb в нисходящем направлении. Это привело к идентификации одного гена-кандидата, AT4G19710 , кодирующего аспартаткиназу-гомосериндегидрогеназу.Этот ген экспрессируется во многих тканях в контрольных условиях (Schmid et al., 2005).
Постантезисная чувствительность к тепловому стрессу
QTL4, расположенный в основании третьей хромосомы, специфичен для теплового ответа в области развития B и, как предполагается, связан с оплодотворением или формированием эмбриона. QTL4 определяется пятью прочно связанными SNP, которые находятся в высоком неравновесном сцеплении (LD> 0,9; дополнительный рисунок 1) с повторно секвенированными SNP в гене, кодирующем кинетохорный белок ( AT3G48210 ). Этот белок участвует в делении клеток — процессе, который имеет решающее значение для растущего эмбриона. Функциональная характеристика этого гена не проводилась, но он экспрессируется во многих тканях, включая эмбрион (Schmid et al., 2005). Высокая степень сцепления (LD> 0,7; дополнительный рисунок 1) также наблюдалась между сильно ассоциированными SNP и повторными последовательностями SNP, расположенными в AT3G48190 и его промоторной области. AT3G48190 , как предполагается, играет роль в репарации ДНК при стрессе (Huefner et al., 2011) и могут быть важны для эмбрионов, подвергшихся тепловому стрессу. Однако медленный распад LD в этой области приводит к блоку сцепления размером 140 т.п.н. (дополнительный рисунок 1), в котором расположены 37 генов, и необходимы дальнейшие анализы для подтверждения причинного гена.
Этот белок участвует в делении клеток — процессе, который имеет решающее значение для растущего эмбриона. Функциональная характеристика этого гена не проводилась, но он экспрессируется во многих тканях, включая эмбрион (Schmid et al., 2005). Высокая степень сцепления (LD> 0,7; дополнительный рисунок 1) также наблюдалась между сильно ассоциированными SNP и повторными последовательностями SNP, расположенными в AT3G48190 и его промоторной области. AT3G48190 , как предполагается, играет роль в репарации ДНК при стрессе (Huefner et al., 2011) и могут быть важны для эмбрионов, подвергшихся тепловому стрессу. Однако медленный распад LD в этой области приводит к блоку сцепления размером 140 т.п.н. (дополнительный рисунок 1), в котором расположены 37 генов, и необходимы дальнейшие анализы для подтверждения причинного гена.
Возможная генетическая связь между временем развития цветения и реакцией на тепловой стресс
Сильно связанный SNP QTL6 на вершине пятой хромосомы находится в LD с 17 SNP, идентифицированными в данных повторной последовательности (дополнительный рисунок 1). Четыре из этих SNP были включены в картирование GWA и были умеренно связаны (-log (значение P)> 4) с ответом на тепловой стресс в области C. Самый сильный ассоциированный SNP отвечает за несинонимичное изменение в последовательности белка AT5G10170, которое коды для мио-инозитол-1-фосфат-синтазы 3 (MIPS3). MIPS3 Экспрессия была наиболее заметной в сосудистой ткани (Donahue et al., 2010), а индуцированная нагреванием экспрессия была обнаружена во время стадии проростков (Khurana et al., 2012). Геном Arabidopsis содержит три гена MIPS .Анализ близкого родственника MIPS2 показал, что экспрессия этого гена сильно индуцируется при воздействии теплового стресса на проростки, цветы и зрелые стручки (Khurana et al., 2012). Исследование теплового ответа у мутанта с нокаутом Т-ДНК MIPS3 не выявило различий между тепловым ответом этой линии и Col-0 дикого типа (дополнительный набор данных 3), что снова указывает на то, что этот ген не может быть причиной наблюдаемая естественная вариация.
Четыре из этих SNP были включены в картирование GWA и были умеренно связаны (-log (значение P)> 4) с ответом на тепловой стресс в области C. Самый сильный ассоциированный SNP отвечает за несинонимичное изменение в последовательности белка AT5G10170, которое коды для мио-инозитол-1-фосфат-синтазы 3 (MIPS3). MIPS3 Экспрессия была наиболее заметной в сосудистой ткани (Donahue et al., 2010), а индуцированная нагреванием экспрессия была обнаружена во время стадии проростков (Khurana et al., 2012). Геном Arabidopsis содержит три гена MIPS .Анализ близкого родственника MIPS2 показал, что экспрессия этого гена сильно индуцируется при воздействии теплового стресса на проростки, цветы и зрелые стручки (Khurana et al., 2012). Исследование теплового ответа у мутанта с нокаутом Т-ДНК MIPS3 не выявило различий между тепловым ответом этой линии и Col-0 дикого типа (дополнительный набор данных 3), что снова указывает на то, что этот ген не может быть причиной наблюдаемая естественная вариация. Кроме того, были проанализированы мутанты двух других генов в области LD QTL6. AT5G10200 кодирует белок, подобный повтору ARM / тетратрикопептидному повтору. Как и у большинства белков ARM-повторов Arabidopsis, функция AT5G10200 неизвестна. У риса полногеномное исследование экспрессии гена повторов ARM показало, что некоторые гены этого семейства регулируются абиотическими стрессами (Sharma et al., 2014). Однако исследование мутанта с нокаутом Т-ДНК не выявило непосредственной роли этого гена в тепловом ответе Arabidopsis (дополнительный набор данных 3). Это согласуется с наблюдением, что другой белок ARM-повтора Arabidopsis, ARO1 , необходим для прорастания пыльцевых трубок и проникновения в рыльце (Gebert et al., 2008). Эти процессы имеют место в постантезе, тогда как QTL6 связан с предантезисными тепловыми реакциями.
Кроме того, были проанализированы мутанты двух других генов в области LD QTL6. AT5G10200 кодирует белок, подобный повтору ARM / тетратрикопептидному повтору. Как и у большинства белков ARM-повторов Arabidopsis, функция AT5G10200 неизвестна. У риса полногеномное исследование экспрессии гена повторов ARM показало, что некоторые гены этого семейства регулируются абиотическими стрессами (Sharma et al., 2014). Однако исследование мутанта с нокаутом Т-ДНК не выявило непосредственной роли этого гена в тепловом ответе Arabidopsis (дополнительный набор данных 3). Это согласуется с наблюдением, что другой белок ARM-повтора Arabidopsis, ARO1 , необходим для прорастания пыльцевых трубок и проникновения в рыльце (Gebert et al., 2008). Эти процессы имеют место в постантезе, тогда как QTL6 связан с предантезисными тепловыми реакциями.
Интересно, что другой ген в LD с QTL6, AT5G10140 , кодирует FLC, который участвует в определении времени цветения. Две изогенные линии, содержащие разные аллели FLC и сильный функциональный аллель FRIGIDA ( FRI ; AT4G00650 ) на фоне Колумбии, были протестированы на их тепловую реакцию. Необходим сильный аллель FRI , потому что FRI является вышестоящим регулятором FLC , и наличие нефункционального аллеля (как в образце дикого типа Columbia) приведет к раннему цветению, независимо от аллеля FLC . .Линия FRI-sf2 , содержащая функциональный колумбийский аллель FLC , требует яровизации для цветения, тогда как линия FRI-flc3 , содержащая нулевой аллель FLC , зацветает рано. Обе линии были яровизированы в течение 10 недель, после чего FRI-sf2 все еще цвел на неделю позже, чем FRI-flc3 . Термическую обработку проводили примерно через 1 неделю после начала цветения. Линия позднего цветения FRI-sf2 была намного более чувствительна к тепловому стрессу в регионах C, D и E, чем линия раннего цветения FRI-flc3 (рис. 6).Обе линии имели аналогичные тепловые отклики для области B, что позволяет предположить, что FLC отвечает за часть теплового отклика до начала реакции.
Необходим сильный аллель FRI , потому что FRI является вышестоящим регулятором FLC , и наличие нефункционального аллеля (как в образце дикого типа Columbia) приведет к раннему цветению, независимо от аллеля FLC . .Линия FRI-sf2 , содержащая функциональный колумбийский аллель FLC , требует яровизации для цветения, тогда как линия FRI-flc3 , содержащая нулевой аллель FLC , зацветает рано. Обе линии были яровизированы в течение 10 недель, после чего FRI-sf2 все еще цвел на неделю позже, чем FRI-flc3 . Термическую обработку проводили примерно через 1 неделю после начала цветения. Линия позднего цветения FRI-sf2 была намного более чувствительна к тепловому стрессу в регионах C, D и E, чем линия раннего цветения FRI-flc3 (рис. 6).Обе линии имели аналогичные тепловые отклики для области B, что позволяет предположить, что FLC отвечает за часть теплового отклика до начала реакции. Этот эксперимент согласуется с наблюдением в ассоциативной панели о том, что тепловая реакция и время цветения отрицательно коррелированы, и предполагает, что эти два процесса, прямо или косвенно, генетически связаны. Чтобы выяснить, является ли этот результат специфичным для аллеля Columbia, шести наиболее распространенных аллелей FLC (P. Li et al., 2014) сравнивали по времени цветения после 10 недель яровизации и по их тепловому отклику. Это сравнение подтвердило отрицательную корреляцию, наблюдаемую между двумя признаками (рис. 7). Образцы с аллелем Col-0 (гаплотип RV2), которые зацвели рано, имели значительно более крупные стебли в области C (рисунок 7) и области D (дополнительная таблица 4), чем образцы, принадлежащие к гаплотипам SV2, SV3 и SV4, которые зацвели позже. . Гаплотип RV1, как и RV2, был идентифицирован из-за его сильной реакции на яровизацию (P.Li et al., 2014), но не было обнаружено значительных различий в длине силиката при нагревании между образцами этого гаплотипа и гаплотипами SV с медленным ответом на яровизацию, что указывает на то, что RV2 представляет собой аллель FLC , связанный с термостойкостью.
Этот эксперимент согласуется с наблюдением в ассоциативной панели о том, что тепловая реакция и время цветения отрицательно коррелированы, и предполагает, что эти два процесса, прямо или косвенно, генетически связаны. Чтобы выяснить, является ли этот результат специфичным для аллеля Columbia, шести наиболее распространенных аллелей FLC (P. Li et al., 2014) сравнивали по времени цветения после 10 недель яровизации и по их тепловому отклику. Это сравнение подтвердило отрицательную корреляцию, наблюдаемую между двумя признаками (рис. 7). Образцы с аллелем Col-0 (гаплотип RV2), которые зацвели рано, имели значительно более крупные стебли в области C (рисунок 7) и области D (дополнительная таблица 4), чем образцы, принадлежащие к гаплотипам SV2, SV3 и SV4, которые зацвели позже. . Гаплотип RV1, как и RV2, был идентифицирован из-за его сильной реакции на яровизацию (P.Li et al., 2014), но не было обнаружено значительных различий в длине силиката при нагревании между образцами этого гаплотипа и гаплотипами SV с медленным ответом на яровизацию, что указывает на то, что RV2 представляет собой аллель FLC , связанный с термостойкостью. Однако между образцами, принадлежащими к одному и тому же гаплотипу, наблюдались большие различия во времени цветения и тепловой реакции, что могло быть следствием вариации последовательности в других локусах времени цветения (Shindo et al., 2005) или генов тепловой реакции.Различия между аллелями больше не были значимыми после поправки на структуру популяции (дополнительный набор данных 4), что соответствует наблюдениям P. Li et al. (2014), что гаплотипы с разным географическим распределением также различались функционально.
Однако между образцами, принадлежащими к одному и тому же гаплотипу, наблюдались большие различия во времени цветения и тепловой реакции, что могло быть следствием вариации последовательности в других локусах времени цветения (Shindo et al., 2005) или генов тепловой реакции.Различия между аллелями больше не были значимыми после поправки на структуру популяции (дополнительный набор данных 4), что соответствует наблюдениям P. Li et al. (2014), что гаплотипы с разным географическим распределением также различались функционально.
Сравнение длины кремния и времени цветения различных гаплотипов FLC.
(A) Длина кремнезема (среднее ± se) в области C при тепловом стрессе.
(B) Время цветения (среднее ± se) после 10 недель яровизации для образцов шести наиболее распространенных гаплотипов FLC (P.Ли и др., 2014). Цифры в столбцах указывают количество образцов, принадлежащих к гаплотипу, указанному под столбцом; Col-0 принадлежит к гаплотипу RV2. Значимые различия (P <0,05), обозначенные буквами над столбиками, определяются с помощью однофакторного дисперсионного анализа с поправкой Бонферрони для множественного тестирования.
Значимые различия (P <0,05), обозначенные буквами над столбиками, определяются с помощью однофакторного дисперсионного анализа с поправкой Бонферрони для множественного тестирования.
Естественная вариация длины кремнистой массы в контрольных условиях указывает на роль аллергенов пыльцы .Для QTL1, контрольного QTL с наиболее сильным ассоциированным SNP и наибольшим размером эффекта, второй, более слабый ассоциированный SNP (-log (значение P) = 4,2) в умеренном LD (LD = 0,424) был обнаружен на 30 т.п.н. ниже по течению. Значимый SNP расположен в интроне гена устойчивости к белой ржавчине (
AT1G56510 ), который высоко экспрессируется в листьях. Более слабый ассоциированный SNP расположен между AT1G56580 , геном, связанным с архитектурой трихома, и AT1G56590 , геном, участвующим в перемещении пузырьков между trans -сетью Гольджи и вакуолями.Ни один из этих трех генов не является очевидным кандидатом. Однако распад LD в этой области был очень медленным, что привело к блоку сцепления почти 176 т. п.н., 8 т.п.н. вверх по течению и 168 т.п.н. ниже значимого SNP (порог: LD = 0,4), в котором расположены 127 генов (дополнительные Рисунок 1). Большинство этих генов являются пре-тРНК (81 ген) или генами с неизвестной биологической функцией (22 гена). Ни один из генов с аннотированной функцией не является очевидным кандидатом, и для определения причинного гена необходимы дальнейшие исследования.Для QTL3 и QTL7 гены-кандидаты могут быть идентифицированы на основе функциональной аннотации. QTL3 определяется двумя значимыми SNP, которые расположены на расстоянии 0,2 т.п.н. друг от друга. Три других умеренно связанных SNP были идентифицированы в непосредственной близости (в пределах 0,4 т.п.н.). Эти пять SNP расположены в промоторе AT3G45970 , на 1,2 т.п.н. выше стартового кодона, и имеют низкую LD (0,2
п.н., 8 т.п.н. вверх по течению и 168 т.п.н. ниже значимого SNP (порог: LD = 0,4), в котором расположены 127 генов (дополнительные Рисунок 1). Большинство этих генов являются пре-тРНК (81 ген) или генами с неизвестной биологической функцией (22 гена). Ни один из генов с аннотированной функцией не является очевидным кандидатом, и для определения причинного гена необходимы дальнейшие исследования.Для QTL3 и QTL7 гены-кандидаты могут быть идентифицированы на основе функциональной аннотации. QTL3 определяется двумя значимыми SNP, которые расположены на расстоянии 0,2 т.п.н. друг от друга. Три других умеренно связанных SNP были идентифицированы в непосредственной близости (в пределах 0,4 т.п.н.). Эти пять SNP расположены в промоторе AT3G45970 , на 1,2 т.п.н. выше стартового кодона, и имеют низкую LD (0,2  , 2002). Значимый SNP QTL7 расположен в большом блоке связи, начиная с 43 kb в восходящем направлении и заканчивая 76 kb в нисходящем направлении от значимого SNP. В этом блоке сцепления были идентифицированы два гена-кандидата: AT5G22430 , ген консервативного семейства аллергенов, который, как предполагается, играет роль во взаимодействиях пыльца-пестик (Jiménez-López et al., 2011), и AT5G22640 , ген, необходимый для правильного развития эмбриона (Liang et al., 2010). Примечательно, что оба идентифицированных экспансиноподобных белка QTL3 и AT5G22430 QTL7 содержат домен аллергена пыльцы.Известно, что белки с таким доменом присутствуют в пыльце и вызывают аллергические реакции у людей. Предполагается, что эти белки ослабляют стенки материнских клеток, чтобы способствовать проникновению стигмы пыльцевой трубкой (Cosgrove, 2000; Шарова, 2007). Несоответствие во взаимодействии пыльцы и пестика может привести к снижению оплодотворения и, следовательно, к более коротким стеблям.
, 2002). Значимый SNP QTL7 расположен в большом блоке связи, начиная с 43 kb в восходящем направлении и заканчивая 76 kb в нисходящем направлении от значимого SNP. В этом блоке сцепления были идентифицированы два гена-кандидата: AT5G22430 , ген консервативного семейства аллергенов, который, как предполагается, играет роль во взаимодействиях пыльца-пестик (Jiménez-López et al., 2011), и AT5G22640 , ген, необходимый для правильного развития эмбриона (Liang et al., 2010). Примечательно, что оба идентифицированных экспансиноподобных белка QTL3 и AT5G22430 QTL7 содержат домен аллергена пыльцы.Известно, что белки с таким доменом присутствуют в пыльце и вызывают аллергические реакции у людей. Предполагается, что эти белки ослабляют стенки материнских клеток, чтобы способствовать проникновению стигмы пыльцевой трубкой (Cosgrove, 2000; Шарова, 2007). Несоответствие во взаимодействии пыльцы и пестика может привести к снижению оплодотворения и, следовательно, к более коротким стеблям. Кроме того, для развития семян важен правильный эмбриогенез. Если этот процесс нарушен, меньше семян будет присутствовать на кремнеземе, что приведет к более коротким кремнеземам.
Кроме того, для развития семян важен правильный эмбриогенез. Если этот процесс нарушен, меньше семян будет присутствовать на кремнеземе, что приведет к более коротким кремнеземам.Наши результаты показывают, что наблюдаемая тепловая реакция зависит как от генотипа, так и от стадии развития цветков и эмбрионов во время тепловой обработки. Картирование GWA привело к идентификации QTL, специфичных для стадии развития, и ковариация между тепловым ответом и временем цветения предполагает прямую или косвенную связь между двумя регуляторными путями. Кроме того, были идентифицированы QTL длины кремния в контрольных условиях. Гены-кандидаты были предложены для большинства QTL, некоторые из которых могут быть предварительно подтверждены путем анализа нокаут-мутантов.
ОБСУЖДЕНИЕ
За последнее десятилетие было приложено много усилий для сбора большого количества естественных вариантов для многих видов, включая модельный вид Arabidopsis. Технологический прогресс позволил детально охарактеризовать генетическое разнообразие в таких коллекциях. Ряд исследований показал использование таких плотно генотипированных природных популяций в подходах к картированию GWA (Cockram et al., 2010; Chan et al., 2011; Chao et al., 2014; Verslues et al., 2014). Такие исследования не только раскрывают большое количество естественных вариаций, присущих видам по многим признакам, но также намекают на гены-кандидаты, являющиеся причиной наблюдаемых вариаций. Последнее является важным аргументом в пользу картирования GWA по сравнению, например, с исследованиями экспрессии генов, поскольку причинные гены часто представляют собой ключевые регуляторы или важные компоненты биологических процессов, тогда как дифференциально экспрессируемые гены могут просто представлять изменения программы транскрипции в ответ на генетические или экологические возмущения.Таким образом, картирование GWA может добавить направленности в генетические регуляторные сети и идентифицировать важные опорные точки в биологических процессах. Помимо генетических нарушений, многие сложные черты могут зависеть от взаимодействия с окружающей средой и сроков развития объекта исследования, осложнение, которым часто пренебрегают.
Ряд исследований показал использование таких плотно генотипированных природных популяций в подходах к картированию GWA (Cockram et al., 2010; Chan et al., 2011; Chao et al., 2014; Verslues et al., 2014). Такие исследования не только раскрывают большое количество естественных вариаций, присущих видам по многим признакам, но также намекают на гены-кандидаты, являющиеся причиной наблюдаемых вариаций. Последнее является важным аргументом в пользу картирования GWA по сравнению, например, с исследованиями экспрессии генов, поскольку причинные гены часто представляют собой ключевые регуляторы или важные компоненты биологических процессов, тогда как дифференциально экспрессируемые гены могут просто представлять изменения программы транскрипции в ответ на генетические или экологические возмущения.Таким образом, картирование GWA может добавить направленности в генетические регуляторные сети и идентифицировать важные опорные точки в биологических процессах. Помимо генетических нарушений, многие сложные черты могут зависеть от взаимодействия с окружающей средой и сроков развития объекта исследования, осложнение, которым часто пренебрегают. Таким образом, включение генотипического разнообразия и зависимости развития в одно исследование нарушения окружающей среды может обеспечить более глубокое понимание процессов развития, участвующих в регуляции реакций на абиотический стресс.
Таким образом, включение генотипического разнообразия и зависимости развития в одно исследование нарушения окружающей среды может обеспечить более глубокое понимание процессов развития, участвующих в регуляции реакций на абиотический стресс.
Определение окна чувствительности развития
В этом исследовании влияние кратковременной жары на фертильность измерялось в зависимости от стадии развития. Наблюдались большие различия в чувствительности различных стадий развития цветков и зародышей к стрессу. Раннее развитие цветков и поздний эмбриогенез, которые, как предполагается, имели место в регионах A и E, соответственно, не были чувствительны к тепловому стрессу у большинства образцов (Рисунок 3, Таблица 1). Были выявлены два минимума средней длины кремнезема, соответствующие уменьшенному завязыванию семян, что указывает на два чувствительных периода в развитии.Один из минимумов, наблюдаемых в нашем эксперименте, соответствует стадиям цветения 9, 10, 11 и 12 (Smyth et al., 1990), представляя стадии развития, на которых происходит мейоз самцов и самок и происходит удлинение пыльников (Таблица 1). Другой минимум соответствует цветкам на стадиях цветения 14 и 15, которые были подвергнуты термообработке после цветения, на этих стадиях происходит раннее формирование зародыша. Мужской мейоз, который происходит на 9 стадии цветения, является процессом, чувствительным к теплу у многих культур (Hedhly, 2011).В более раннем исследовании Arabidopsis изучалось влияние тепла на развитие цветов (но не на развитие зародыша) у образца Columbia путем подсчета плодородных стручков вдоль основного соцветия. Хотя термическая обработка в этом исследовании была более жесткой (42 ° C / 4 ч), наблюдался чувствительный период на стадии 9 цветения, как и в нашем исследовании. Кроме того, был выявлен чувствительный период на стадии цветения 12 (Kim et al., 2001). Более обширное исследование Arabidopsis, изучающее прерывание цветковых почек при тепловом стрессе у 13 естественных образцов, показало, что у образцов Columbia цветочная стадия 12, но не цветочная стадия 9, была наиболее чувствительной (Warner and Erwin, 2005).
Другой минимум соответствует цветкам на стадиях цветения 14 и 15, которые были подвергнуты термообработке после цветения, на этих стадиях происходит раннее формирование зародыша. Мужской мейоз, который происходит на 9 стадии цветения, является процессом, чувствительным к теплу у многих культур (Hedhly, 2011).В более раннем исследовании Arabidopsis изучалось влияние тепла на развитие цветов (но не на развитие зародыша) у образца Columbia путем подсчета плодородных стручков вдоль основного соцветия. Хотя термическая обработка в этом исследовании была более жесткой (42 ° C / 4 ч), наблюдался чувствительный период на стадии 9 цветения, как и в нашем исследовании. Кроме того, был выявлен чувствительный период на стадии цветения 12 (Kim et al., 2001). Более обширное исследование Arabidopsis, изучающее прерывание цветковых почек при тепловом стрессе у 13 естественных образцов, показало, что у образцов Columbia цветочная стадия 12, но не цветочная стадия 9, была наиболее чувствительной (Warner and Erwin, 2005). Эта цветочная стадия 9 не была идентифицирована, что могло быть вызвано более длительным периодом воздействия тепла (36 ° C в течение 72 часов). Однако было показано, что другие образцы более чувствительны, чем Columbia, и прерывание цветковых бутонов также наблюдалось на более ранних стадиях, чем стадия 12 цветков. Стадия 12 цветков соответствует удлинению пыльников. Снижение удлинения пыльников наблюдалось при тепловом стрессе в результате снижения уровня ауксина в развивающихся пыльниках. Уменьшение удлинения пыльников при тепловом стрессе также было обнаружено у других видов, таких как ячмень (Sakata et al., 2010). Наблюдения Кима и др. (2001) и Warner and Erwin (2005) согласуются с нашим наблюдением, что естественные вариации существуют как с точки зрения продолжительности окна чувствительности, так и степени уменьшения количества семян, установленных в пределах окна чувствительности. Тем не менее, сравнение стадий цветения в различных исследованиях следует проводить осторожно, поскольку соответствие между областями развития и знаковыми событиями в развитии цветков и семян основано на обобщении, и наблюдались естественные вариации количества цветков, раскрывающихся за день.
Эта цветочная стадия 9 не была идентифицирована, что могло быть вызвано более длительным периодом воздействия тепла (36 ° C в течение 72 часов). Однако было показано, что другие образцы более чувствительны, чем Columbia, и прерывание цветковых бутонов также наблюдалось на более ранних стадиях, чем стадия 12 цветков. Стадия 12 цветков соответствует удлинению пыльников. Снижение удлинения пыльников наблюдалось при тепловом стрессе в результате снижения уровня ауксина в развивающихся пыльниках. Уменьшение удлинения пыльников при тепловом стрессе также было обнаружено у других видов, таких как ячмень (Sakata et al., 2010). Наблюдения Кима и др. (2001) и Warner and Erwin (2005) согласуются с нашим наблюдением, что естественные вариации существуют как с точки зрения продолжительности окна чувствительности, так и степени уменьшения количества семян, установленных в пределах окна чувствительности. Тем не менее, сравнение стадий цветения в различных исследованиях следует проводить осторожно, поскольку соответствие между областями развития и знаковыми событиями в развитии цветков и семян основано на обобщении, и наблюдались естественные вариации количества цветков, раскрывающихся за день.
Также у других видов Brassicaceae сообщалось о тепловом отклике, аналогичном нашим наблюдениям. Исследования видов рапса и горчицы ( B. napus и B. juncea ) показали, что тепловой стресс во время цветения или развития стручков приводит к большему снижению урожайности, чем тепловой стресс во время формирования бутонов (Gan et al., 2004). Кроме того, было замечено, что стручки, прошедшие критический порог развития, были толерантны к тепловому стрессу (Angadi et al., 2000). Этот порог соответствует позднему эмбриогенезу, стадии развития, которая также не была чувствительна к теплу ни у одного образца, протестированного в нашем эксперименте. Несмотря на то, что исследование Angadi et al. (2000) подтверждают существование окна чувствительности развития, в котором тепловой стресс влияет на урожайность, сравнение этих результатов с наблюдениями в нашем эксперименте также обнаруживает интересную разницу. Мы наблюдали более сильное воздействие стресса перед цветением, в то время как у других Brassicaceae воздействие, по-видимому, более преобладающее после начала цветения. Это может быть связано с абортом эмбриона во время раннего развития кремнезема, что наблюдалось в наших экспериментах (Рисунок 1). Более того, результаты Gan et al. (2004) основаны на общем урожае семян и требуют осторожного толкования. Если механизмы компенсации активны (Рисунок 4; Angadi et al., 2003), общий урожай семян может быть не лучшим показателем для определения окна чувствительности развития.
Это может быть связано с абортом эмбриона во время раннего развития кремнезема, что наблюдалось в наших экспериментах (Рисунок 1). Более того, результаты Gan et al. (2004) основаны на общем урожае семян и требуют осторожного толкования. Если механизмы компенсации активны (Рисунок 4; Angadi et al., 2003), общий урожай семян может быть не лучшим показателем для определения окна чувствительности развития.
У видов, отличных от Brassicaceae , сообщалось о тепловых реакциях, подобных нашим наблюдениям.У зеленой фасоли ( Phaseolus vulgaris ) наиболее чувствительный период был за 10 дней до цветения и мог быть связан со снижением жизнеспособности пыльцы (Suzuki et al., 2001). Это указывает на связь с мужским мейозом, что соответствует чувствительной стадии 9 цветка у Arabidopsis. У томатов бутоны цветков за 9–5 дней до цветения и цветы через 1–3 дня после цветения очень чувствительны к высоким температурам (Sugiyama et al., 1965). Эти два периода соответствуют двум минимумам, выявленным в нашем эксперименте.
Обнаружение того, что наши наблюдения за арабидопсисом согласуются с данными, собранными для тепловых реакций у различных культур, увеличивает потенциал перевода наших результатов о молекулярной регуляции теплового ответа на виды сельскохозяйственных культур. Интересно, что помимо теплового стресса засуха также влияет на развитие цветов и эмбрионов. У зерновых есть два чувствительных периода при стрессе засухи: первый сосредоточен вокруг периода мужского и женского мейоза, а второй — во время цветения и начального развития зерна (Saini, 1997).Это говорит о том, что могут быть задействованы аналогичные регуляторные механизмы, что увеличивает актуальность этой работы.
GWA-картирование выявляет специфичные для стадии QTL
Хотя в нескольких исследованиях сообщалось об окне чувствительности развития при тепловом стрессе (Kim et al., 2001; Warner and Erwin, 2005), это исследование является уникальным, поскольку оно исследует это окно чувствительности. используя большую естественную популяцию. Следовательно, данные могут быть подвергнуты картированию GWA, что позволило идентифицировать специфичные для стадии QTL.Два варианта в нашей экспериментальной установке имели решающее значение для определения QTL, специфичных для стадии. Во-первых, 1-дневная выдержка гарантировала, что развивающиеся цветы подверглись стрессу только на одной стадии развития. В экспериментах, в которых стресс применяется в течение более длительных периодов времени, идентификация специфичных для стадии QTL менее точна, потому что один цветок может испытывать стресс на нескольких стадиях развития. Во-вторых, количественная оценка воздействия теплового стресса путем измерения длины отдельных стручков вдоль основного соцветия обеспечила высокое разрешение тепловых эффектов, связанных с развитием.Исследования, которые количественно оценивают реакцию на тепловой стресс по общему урожаю семян или фруктов, будут иметь меньше возможностей для выявления эффектов, которые присутствуют только в небольшом окне развития.
Следовательно, данные могут быть подвергнуты картированию GWA, что позволило идентифицировать специфичные для стадии QTL.Два варианта в нашей экспериментальной установке имели решающее значение для определения QTL, специфичных для стадии. Во-первых, 1-дневная выдержка гарантировала, что развивающиеся цветы подверглись стрессу только на одной стадии развития. В экспериментах, в которых стресс применяется в течение более длительных периодов времени, идентификация специфичных для стадии QTL менее точна, потому что один цветок может испытывать стресс на нескольких стадиях развития. Во-вторых, количественная оценка воздействия теплового стресса путем измерения длины отдельных стручков вдоль основного соцветия обеспечила высокое разрешение тепловых эффектов, связанных с развитием.Исследования, которые количественно оценивают реакцию на тепловой стресс по общему урожаю семян или фруктов, будут иметь меньше возможностей для выявления эффектов, которые присутствуют только в небольшом окне развития. Механизмы компенсации растений (Рисунок 4; Angadi et al., 2003; Young et al., 2004) делают общий урожай семян или плодов еще менее надежным. Наши результаты должны повысить осведомленность о том факте, что регуляция интересующей черты может изменяться в процессе развития. Дизайн экспериментов является здесь решающим фактором, который в большинстве случаев определяет, можно ли обнаружить специфичность стадии развития QTL или регуляторных сетей или нет.Точные измерения или большие размеры популяции не будут способствовать более высокой степени обнаружения QTL, специфичных для стадии, когда выбран неоптимальный дизайн.
Механизмы компенсации растений (Рисунок 4; Angadi et al., 2003; Young et al., 2004) делают общий урожай семян или плодов еще менее надежным. Наши результаты должны повысить осведомленность о том факте, что регуляция интересующей черты может изменяться в процессе развития. Дизайн экспериментов является здесь решающим фактором, который в большинстве случаев определяет, можно ли обнаружить специфичность стадии развития QTL или регуляторных сетей или нет.Точные измерения или большие размеры популяции не будут способствовать более высокой степени обнаружения QTL, специфичных для стадии, когда выбран неоптимальный дизайн.
В нашем исследовании основное внимание уделялось идентификации специфичных для стадии QTL. Два минимума были идентифицированы в длине кремнезема после теплового стресса, соответствующие двум отдельным процессам в развитии цветов и семян (Рисунок 3, Таблица 1). Информация по этапам может быть полезна для определения приоритетов генов-кандидатов для будущих функциональных анализов.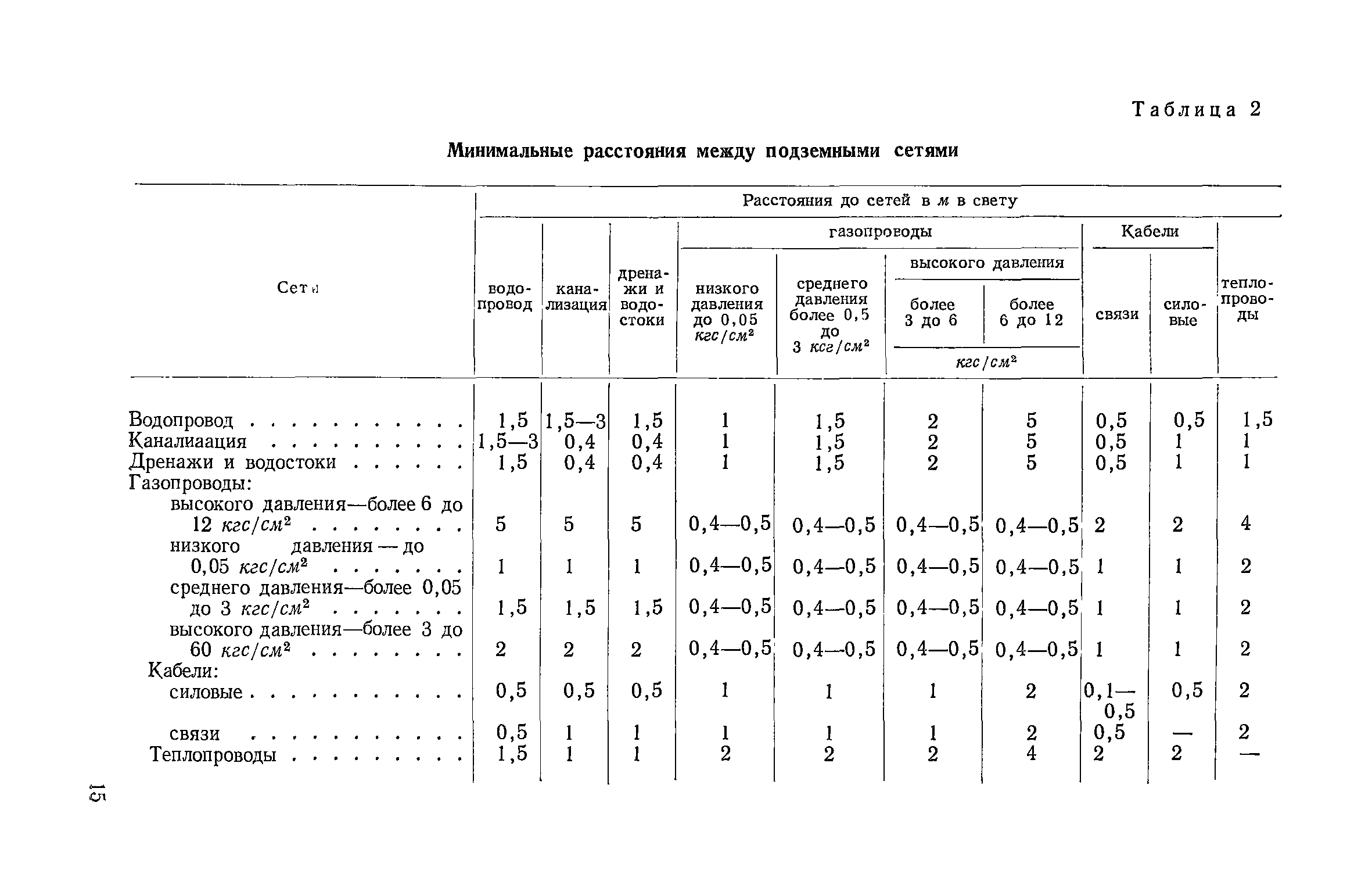 Гены, которые не экспрессируются на стадии развития, на которой идентифицирован QTL, или которые не экспрессируются в цветках или эмбрионах, могут иметь более низкий приоритет. В этом исследовании четыре QTL были идентифицированы как сильно связанные с тепловым ответом: один специфичный для ответа после цветения и три специфических для ответа до цветения. Основываясь на LD, аннотированной функции гена, профилях экспрессии и анализе мутантов, один или два гена на QTL могут быть приоритетными в качестве генов-кандидатов. Нокаут-анализ нескольких генов-кандидатов показал участие этих генов в ответах репродуктивных процессов на тепловой стресс.Однако для ряда генов их участие не удалось подтвердить. Это может быть связано с генетическим фоном мутантной линии (Col-0), дикий тип которой используется для сравнения. С другой стороны, нокауты, которые действительно показывают значительное влияние на изучаемый признак, не исключают окончательно участие других генов, объясняющих наблюдаемые вариации в окне поддержки QTL.
Гены, которые не экспрессируются на стадии развития, на которой идентифицирован QTL, или которые не экспрессируются в цветках или эмбрионах, могут иметь более низкий приоритет. В этом исследовании четыре QTL были идентифицированы как сильно связанные с тепловым ответом: один специфичный для ответа после цветения и три специфических для ответа до цветения. Основываясь на LD, аннотированной функции гена, профилях экспрессии и анализе мутантов, один или два гена на QTL могут быть приоритетными в качестве генов-кандидатов. Нокаут-анализ нескольких генов-кандидатов показал участие этих генов в ответах репродуктивных процессов на тепловой стресс.Однако для ряда генов их участие не удалось подтвердить. Это может быть связано с генетическим фоном мутантной линии (Col-0), дикий тип которой используется для сравнения. С другой стороны, нокауты, которые действительно показывают значительное влияние на изучаемый признак, не исключают окончательно участие других генов, объясняющих наблюдаемые вариации в окне поддержки QTL.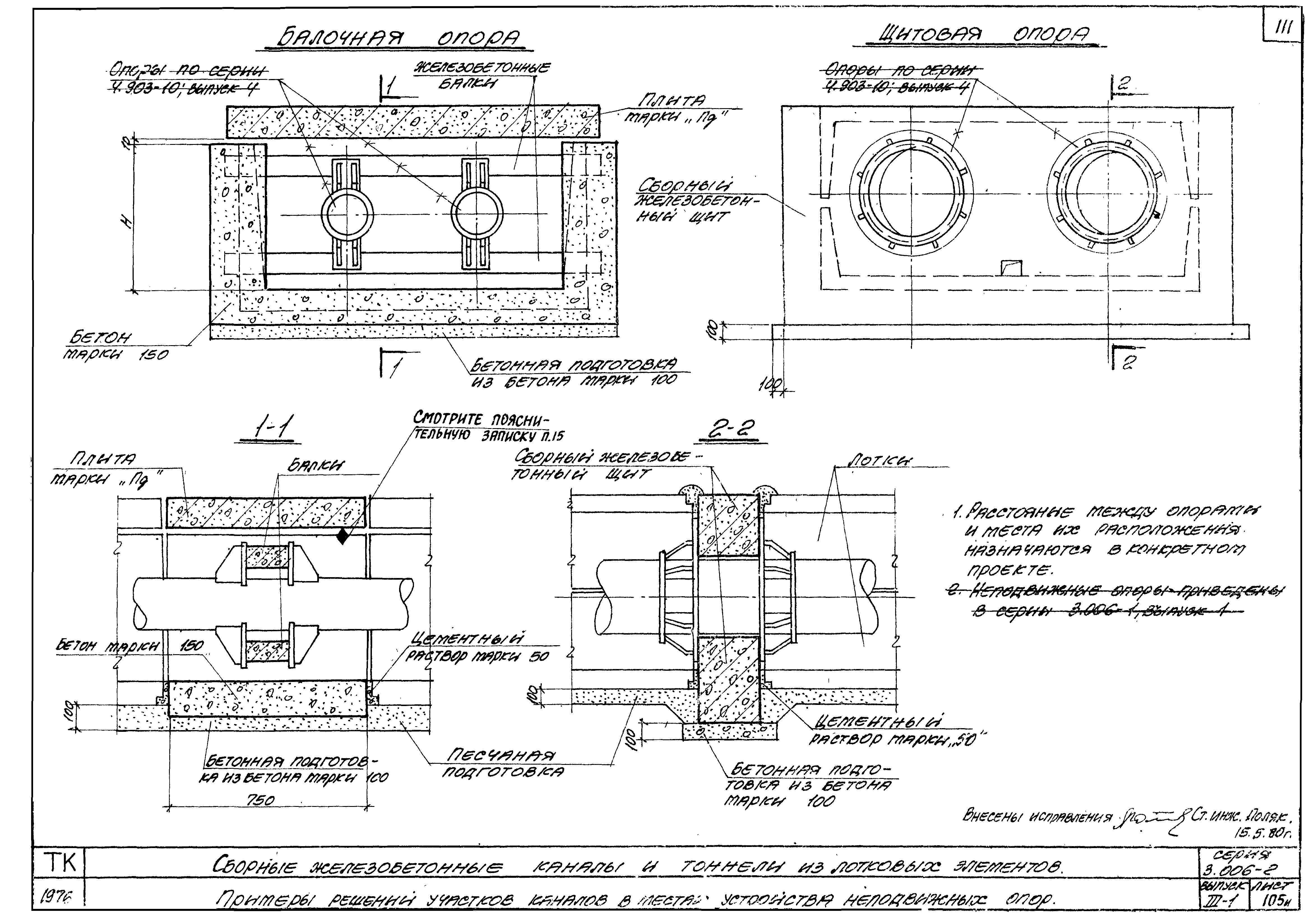 Дальнейшие доказательства должны быть получены из исследований функциональной комплементации в различных генетических фонах.
Дальнейшие доказательства должны быть получены из исследований функциональной комплементации в различных генетических фонах.
Во многих исследованиях, описывающих тепловой стресс во время цветения, видная роль отводится белкам теплового шока (Hsps) и факторам транскрипции теплового шока (Hsfs) (Giorno et al., 2013). Напр., Конститутивное и вызванное тепловым стрессом накопление Hsps в репродуктивной ткани (особенно в пыльниках) связано с термостойкостью (Bita et al., 2011). Hsps стабилизируют белки, которые разворачиваются из-за теплового стресса (Al-Whaibi, 2011). Hsps, как шапероны, предотвращают необратимую агрегацию других белков и участвуют в рефолдинге белков в условиях теплового стресса (Waters, 2013).Удивительно, но в нашем исследовании не было обнаружено Hsp или Hsf в непосредственной близости от QTL (порог –log (P) = 5,5), и только два Hsps были идентифицированы в пределах 20 т.п.н. от умеренно ассоциированных SNP (пороговое значение –log (P) = 4), предполагая, что аллельные вариации Hsps или Hsfs не являются основной причиной естественных вариаций термостойкости во время цветения. Отсутствие этих очевидных кандидатов в наших результатах показывает, что нецелевой анализ, такой как картирование GWA, во многих случаях приводит к идентификации новых функций генов, в то время как исследования, основанные на гипотезах, в большинстве случаев приводят к доказательству или опровержению. уже предложенной функции.
Отсутствие этих очевидных кандидатов в наших результатах показывает, что нецелевой анализ, такой как картирование GWA, во многих случаях приводит к идентификации новых функций генов, в то время как исследования, основанные на гипотезах, в большинстве случаев приводят к доказательству или опровержению. уже предложенной функции.
Генетическая регуляция теплового ответа и цветения
Высокая отрицательная корреляция и совпадение сильно (таблица 4) и умеренно связанных SNP (дополнительный набор данных 2) наблюдались для устойчивости к тепловому стрессу и времени цветения. QTL6 был определен для реакции на тепловой стресс, но также умеренно связан со временем цветения. Значимый SNP QTL6 был связан с SNP в FLC , репрессоре времени цветения, и его промоторе. Влияние теплового стресса оценивалось на двух изогенных линиях: одна с функциональным аллелем FLC , требующая яровизации для цветения, и вторая, содержащая нулевой аллель FLC , приводящая к раннему цветению. Тепловой стресс привел к значительному сокращению длины стручка в позднецветущей линии по сравнению с раннецветущей. Анализ шести наиболее распространенных гаплотипов FLC дополнительно подтвердил, что тепловая чувствительность связана с поздним цветением и может быть основана на перекрестных помехах между регулирующими сетями. Несмотря на то, что изменение реакции на тепловой стресс сопоставлено с FLC , мы не можем исключить косвенное влияние разницы во времени цветения. Особенно потому, что экспрессия FLC снижается в течение 10 недель яровизации, серьезно снижая вариабельность экспрессии FLC во время лечения (Michaels and Amasino, 1999; Shindo et al., 2006). Хотя другие известные генетические факторы, влияющие на время цветения, также могут косвенно способствовать изменению реакции на тепловой стресс, анализ этой взаимосвязи выходит за рамки данного исследования. Положительная корреляция была также обнаружена для устойчивости к холоду и времени цветения у сои, где высокая устойчивость была связана с поздним цветением (Kurosaki et al.
Тепловой стресс привел к значительному сокращению длины стручка в позднецветущей линии по сравнению с раннецветущей. Анализ шести наиболее распространенных гаплотипов FLC дополнительно подтвердил, что тепловая чувствительность связана с поздним цветением и может быть основана на перекрестных помехах между регулирующими сетями. Несмотря на то, что изменение реакции на тепловой стресс сопоставлено с FLC , мы не можем исключить косвенное влияние разницы во времени цветения. Особенно потому, что экспрессия FLC снижается в течение 10 недель яровизации, серьезно снижая вариабельность экспрессии FLC во время лечения (Michaels and Amasino, 1999; Shindo et al., 2006). Хотя другие известные генетические факторы, влияющие на время цветения, также могут косвенно способствовать изменению реакции на тепловой стресс, анализ этой взаимосвязи выходит за рамки данного исследования. Положительная корреляция была также обнаружена для устойчивости к холоду и времени цветения у сои, где высокая устойчивость была связана с поздним цветением (Kurosaki et al. , 2004). Было показано, что различия в толерантности связаны с естественными вариациями в хорошо известном локусе созревания. Хотя реакция на неоптимальные температуры может быть связана с естественным изменением времени цветения или генов созревания, мы не можем исключить, что корреляция, обнаруженная в нашем эксперименте, была вызвана холодным грунтованием.Время между холодным периодом, необходимым для синхронизации цветения, и обработкой не одинаково для всех образцов: образцы, которые зацвели раньше, по сути, получали обработку раньше после обработки холодом, чем позже цветущие. Возможно, растения, недавно обработанные холодной грунтовкой, менее чувствительны к тепловому стрессу. Это грунтовка может высвобождаться со временем, что со временем приводит к повышенной чувствительности к тепловому стрессу. Однако для образцов, подвергшихся термообработке в тот же день после обработки холодом, наблюдалась естественная изменчивость, что указывает на то, что большая часть естественной изменчивости не может быть объяснена различиями в периоде между холодной и термической обработкой.
, 2004). Было показано, что различия в толерантности связаны с естественными вариациями в хорошо известном локусе созревания. Хотя реакция на неоптимальные температуры может быть связана с естественным изменением времени цветения или генов созревания, мы не можем исключить, что корреляция, обнаруженная в нашем эксперименте, была вызвана холодным грунтованием.Время между холодным периодом, необходимым для синхронизации цветения, и обработкой не одинаково для всех образцов: образцы, которые зацвели раньше, по сути, получали обработку раньше после обработки холодом, чем позже цветущие. Возможно, растения, недавно обработанные холодной грунтовкой, менее чувствительны к тепловому стрессу. Это грунтовка может высвобождаться со временем, что со временем приводит к повышенной чувствительности к тепловому стрессу. Однако для образцов, подвергшихся термообработке в тот же день после обработки холодом, наблюдалась естественная изменчивость, что указывает на то, что большая часть естественной изменчивости не может быть объяснена различиями в периоде между холодной и термической обработкой.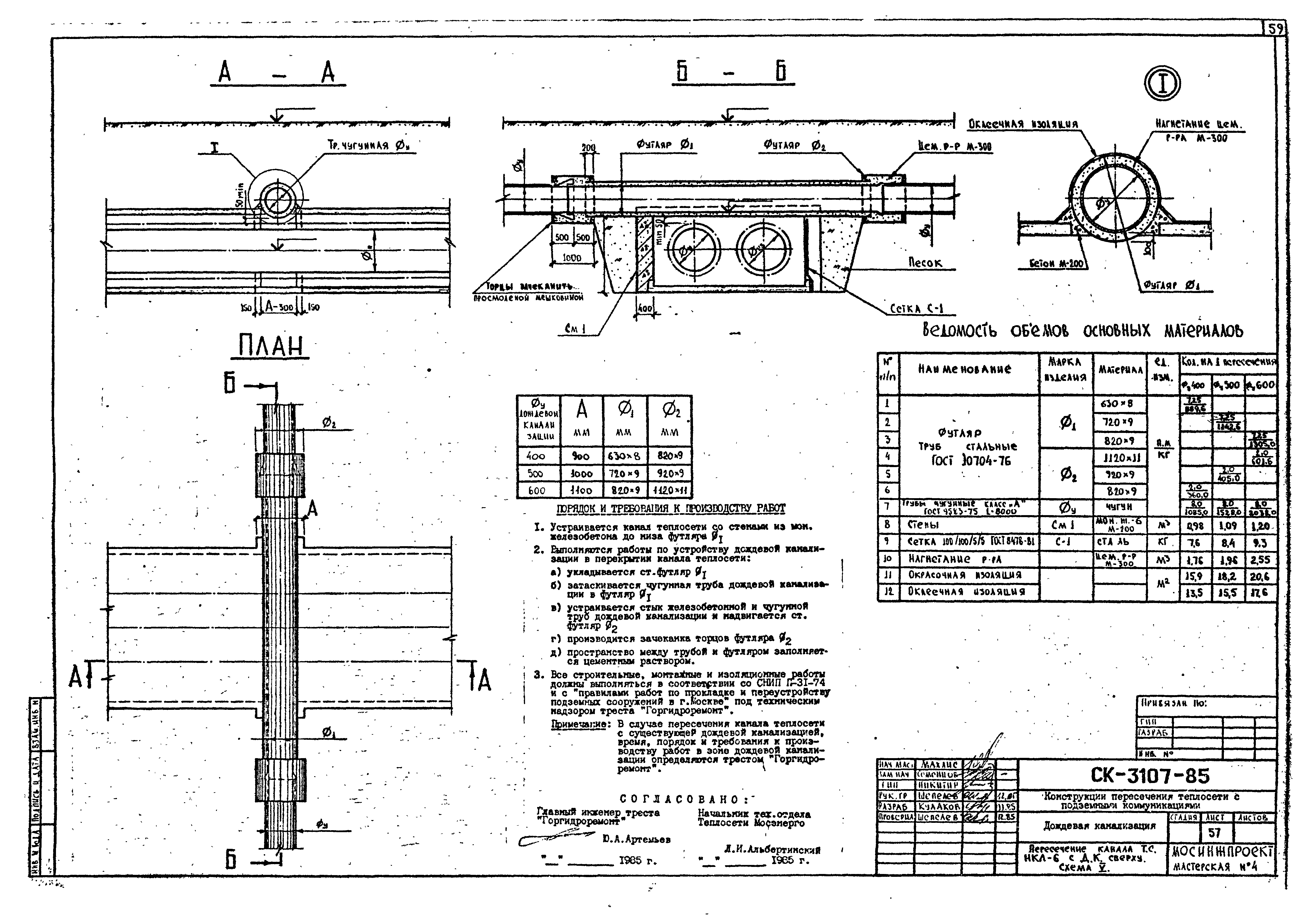 Таким образом, мы предполагаем, что различия в термостойкости, наблюдаемые между ранними и позднецветущими образцами, являются результатом местной адаптации. Раннецветущие образцы — это летние однолетние растения, которые прорастают весной и цветут летом, тогда как позднецветущие образцы — это зимние однолетние растения, которые прорастают осенью, переживают зиму на вегетативной стадии и цветут весной. Это говорит о том, что раноцветущие образцы выиграют от механизма термостойкости своих цветов, тогда как позднецветущие — нет, потому что цветы редко будут подвергаться воздействию высоких температур весной.
Таким образом, мы предполагаем, что различия в термостойкости, наблюдаемые между ранними и позднецветущими образцами, являются результатом местной адаптации. Раннецветущие образцы — это летние однолетние растения, которые прорастают весной и цветут летом, тогда как позднецветущие образцы — это зимние однолетние растения, которые прорастают осенью, переживают зиму на вегетативной стадии и цветут весной. Это говорит о том, что раноцветущие образцы выиграют от механизма термостойкости своих цветов, тогда как позднецветущие — нет, потому что цветы редко будут подвергаться воздействию высоких температур весной.
Гомология реакций на тепловой стресс между видами
Как уже упоминалось, у нескольких видов сельскохозяйственных культур наблюдаются сходные стадийно-специфические тепловые реакции, что предполагает общие регуляторные механизмы реакции растений на повышенную температуру во время цветения, а также на завязывание плодов и семян. Такой общий ответный механизм является многообещающим для трансляции результатов от модельных видов к сельскохозяйственным культурам.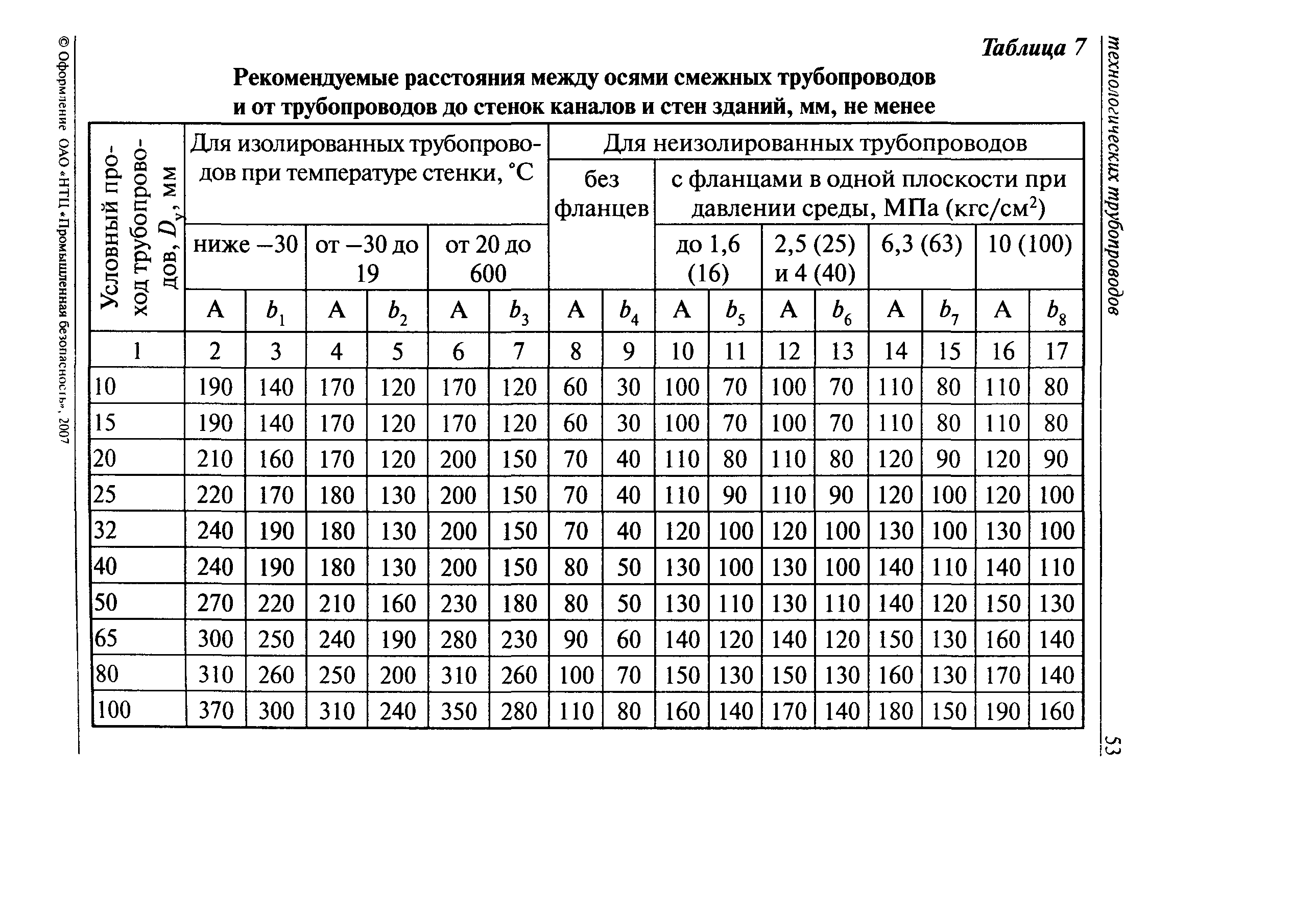 К сожалению, сравнение QTL, наблюдаемых в этом исследовании, с ранее идентифицированными QTL в других исследованиях по картированию сельскохозяйственных культур, направленных на идентификацию генов термостойкости во время цветения, затруднено, поскольку большинство QTL еще не были клонированы (Grilli et al., 2007; Ye et al., 2012; Лукас и др., 2013). Все более ранние исследования проводились на популяции двух родителей с низким разрешением; поэтому связанные области все еще содержат много генов. Поскольку ни одна из протестированных культур (рис, томат и вейный горох) близко не родственна Arabidopsis, сравнение участков генома затруднено из-за отсутствия синтении. Однако из-за высокого разрешения картирования GWA и огромного количества предварительной информации, доступной по Arabidopsis, многообещающе проследить за приоритетными генами-кандидатами, идентифицированными в этом исследовании, с дальнейшим функциональным анализом, чтобы идентифицировать их функции и впоследствии иметь возможность идентифицировать тепловая сеть.
К сожалению, сравнение QTL, наблюдаемых в этом исследовании, с ранее идентифицированными QTL в других исследованиях по картированию сельскохозяйственных культур, направленных на идентификацию генов термостойкости во время цветения, затруднено, поскольку большинство QTL еще не были клонированы (Grilli et al., 2007; Ye et al., 2012; Лукас и др., 2013). Все более ранние исследования проводились на популяции двух родителей с низким разрешением; поэтому связанные области все еще содержат много генов. Поскольку ни одна из протестированных культур (рис, томат и вейный горох) близко не родственна Arabidopsis, сравнение участков генома затруднено из-за отсутствия синтении. Однако из-за высокого разрешения картирования GWA и огромного количества предварительной информации, доступной по Arabidopsis, многообещающе проследить за приоритетными генами-кандидатами, идентифицированными в этом исследовании, с дальнейшим функциональным анализом, чтобы идентифицировать их функции и впоследствии иметь возможность идентифицировать тепловая сеть. Трансляция сети ответов от одного вида к другому может быть более плодотворной, чем сравнение отдельных генов (Ferrier et al., 2011). Гомологи белков в этом пути могут быть идентифицированы, а знания о функции и экспрессии белков, взаимодействиях белок-белок и взаимодействиях белок-метаболит могут быть использованы для идентификации аналогичного пути ответа у других видов (Assmann, 2013).
Трансляция сети ответов от одного вида к другому может быть более плодотворной, чем сравнение отдельных генов (Ferrier et al., 2011). Гомологи белков в этом пути могут быть идентифицированы, а знания о функции и экспрессии белков, взаимодействиях белок-белок и взаимодействиях белок-метаболит могут быть использованы для идентификации аналогичного пути ответа у других видов (Assmann, 2013).
Хотя в данной статье основное внимание уделяется длине кремнезема у растений, подвергшихся тепловому стрессу, результаты, полученные от растений, выращенных в контрольных условиях, также актуальны для целей селекции.Для повышения урожайности и, следовательно, получения большего количества масла, интенсивно ведется селекция сортов B. napus и B. juncea . Общий урожай определяется несколькими характеристиками растения, такими как общее количество плодов на растение, масса семян, количество семян на плод и длина плодов. Последние три коррелированы, и сообщалось о многих совпадающих QTL (Alonso-Blanco et al., 1999; Dechaine et al., 2014; N. Li et al., 2014). Длина плода считается подходящим признаком для выбора в B.napus (Samizadeh et al., 2010), поскольку по этому признаку наблюдается высокая наследуемость, которая менее чувствительна к GxE, чем, например, общее количество плодов на растении (Chay, Thurling, 1989; Dechaine et al., 2014 ). В этом эксперименте высокая наследуемость наблюдалась также по длине кремния (таблица 3). Поскольку Arabidopsis является членом семейства Brassicaceae , аналогичные механизмы, вероятно, будут определять длину стручка, и перенос результатов картирования с этого модельного вида на культуры должен быть относительно простым.
Три гена-кандидата были идентифицированы по длине силикатной оболочки в контрольных условиях, и все они содержали домен аллергена пыльцы. Известно, что аллергены на пыльцу вызывают у людей аллергические реакции, поэтому они интенсивно изучаются. Хотя их биологическая функция у растений еще полностью не выяснена (Jiménez-López et al., 2011), предполагается, что эти белки могут ослаблять клеточную стенку материнской ткани, чтобы облегчить проникновение пыльцевой трубки в рыльце (Cosgrove и другие., 1997; Mollet et al., 2013). На томатах были проведены эксперименты по конкуренции между пыльцой дикого типа и пыльцой, в которой один из этих аллергенов пыльцы был мутирован. Было обнаружено, что пыльца с мутировавшим белком уступает место пыльце дикого типа из-за более медленного роста пыльцевых трубок, что указывает на важную роль пыльцевых аллергенов в доступе пыльцы к яйцеклетке (Valdivia et al., 2007). Длина кремнезема коррелирует с количеством семян на кремнезем (рис. 1). Более медленное проникновение в пыльцевую трубку может привести к менее эффективному оплодотворению; как следствие, разовьется меньше семян и стерни будут короче.По клиническим иммунологическим причинам вариабельность этих аллергенов изучается между видами и внутри видов (Radauer and Breiteneder, 2006; Songnuan, 2013), но влияние естественных вариаций на успех оплодотворения еще не изучено.
В заключение, выбранный экспериментальный дизайн был очень подходящим для определения генетических регуляторов теплового ответа репродуктивных процессов, специфичных для стадии развития. Предварительная проверка нокаут-анализом была предоставлена для двух предварительных QTL, что усилило кандидатуру двух генов в непосредственной близости от значительно ассоциированных SNP.Кроме того, высокое разрешение картирования GWA, сходство между развитием цветов и семян у разных видов, а также предполагаемые общие регуляторные механизмы теплового ответа делают перевод результатов, полученных на арабидопсисе, на культуры многообещающими. Для достижения этой цели необходимы дальнейшие функциональные анализы генов-кандидатов у Arabidopsis и их гомологов в сельскохозяйственных культурах.
МЕТОДЫ
Растительный материал
Коллекция, состоящая из 285 природных образцов Arabidopsis thaliana (дополнительный набор данных 1), отобранных для отражения большей части генетических вариаций, присутствующих внутри вида (Platt et al., 2010) и генотипированный по ~ 215 тыс. Маркеров SNP (Affymetrix SNP-tilling array Atsnptile1; Atwell et al., 2010; Li et al., 2010) был получен из ABRC Stock Center (Baxter et al., 2010).
Для функционального генного анализа две линии, FRI-sf2 и FRI-flc3 , содержащие два разных аллеля FLC на фоне Колумбии (Michaels and Amasino, 1999), были любезно предоставлены M. Koornneef ( MPI Köln). FRI-sf2 (CS6209) также можно получить на складе ABRC.Кроме того, следующие мутанты по вставке Т-ДНК SALK (Alonso et al., 2003) и мутанты по вставке Т-ДНК GABI-Kat (Kleinboelting et al., 2012) были получены из NASC: qul2-2 (SALK_065604c), AT2G03505 (GABI_745A07), AT2G03490 (SALK_125910), mips3-2 (SALK_120131c; Донахью и др., 2010) и AT5G10200 (SALcK_0409). Семена мутантов qul1-1 и qul1-1 qul2-1 любезно предоставлены Л. Остергаардом (Fuentes et al., 2010). Растения, несущие вставку гомозиготной Т-ДНК, генотипировали с помощью ПЦР с использованием праймеров, перечисленных в дополнительной таблице 1.
Экспериментальная установка
Для каждого образца восемь растений выращивали в контролируемых условиях. Пять повторов подвергались термической обработке, а три повтора служили контролем. Семена высевали в чашки Петри на влажную фильтровальную бумагу. После 4 дней обработки холодом их помещали на свет при комнатной температуре на полтора дня для прорастания. Проросшие семена помещали на блоки из минеральной ваты, насыщенные питательным раствором (Hyponex, 1 мМ N, 1.1 мМ P и 5,9 мМ K) в климатической комнате (16 часов света, 125 мкмоль / с / м 2 , влажность 70%, 20/18 ° C). Три раза в неделю блоки минеральной ваты пропитывались питательным раствором с помощью автоматической системы затопления. Через 3 недели растения яровировали в течение 10 недель для синхронизации цветения. После яровизации растения возвращали в климатическую комнату (условия, указанные выше). Через одну-две недели после того, как первая реплика образца начала цвести, ко всем репликам этого образца была применена термообработка.Небольшое количество образцов подверглось обработке за пределами этого окна из-за большой разницы во времени цветения между повторностями (дополнительный набор данных 1). Первый распустившийся в день обработки цветок был помечен ниткой. Три реплики на образец хранились в климатической комнате в качестве контрольных. По пять экземпляров каждого образца переносили в климатический шкаф, где они подвергались термообработке. В начале дня температура повышалась с 20 до 35 ° С за 2 ч.Температуру поддерживали при 35 ° C в течение 13,5 ч. По окончании светового периода снова понижали температуру до 20 ° С на 2 ч. На следующий день после обработки растения возвращали в климатическую комнату.
Эту же процедуру использовали для проверки теплового отклика в FRI-sf2 и FRI-flc3. Для других мутантов со вставкой Т-ДНК, использованных в этом исследовании, была использована та же процедура, но без периода яровизации. Все линии были протестированы в двух независимых экспериментах (эксперименты A и B).
Сбор и анализ основных соцветий Siliques
Когда растение перестало давать новые цветы или когда по крайней мере 40 цветков над меткой нитки превратились в стручки, основное соцветие было отрезано и сохранено в бумажном пакете для последующего анализа.Контрольные и обработанные растения одного и того же образца всегда собирали в один и тот же день. Сохраненный материал использовали для определения длины всех стручков, присутствующих в основном соцветии. Кремний, помеченный нитью, был помечен как позиция номер 0. Siliques под тегом получил отрицательные номера позиций, а Siliques над тегом получил положительные номера позиций (рис. 2).
Свойства семян
Чтобы определить количество семян на кремневую стружку и их размер, семена были распределены на белой бумаге, и фотографии были сделаны в условиях постоянного освещения, чтобы облегчить обработку изображений.Снимки обрабатывались с помощью программного пакета ImageJ (Schneider et al., 2012). На основе распределения по размеру темно-коричневых и светло-коричневых семян порог 0,12 мм 2 был использован для определения семян малого и нормального размера, что указывает на нежизнеспособные и жизнеспособные семена, соответственно. Для определения всхожести семена помещали на влажную фильтровальную бумагу в чашки Петри, запечатанные Parafilm. Чашки Петри помещали в темноту при 4 ° C на 4 дня. После этого чашки Петри помещали на свет при комнатной температуре.Подсчет количества проросших и непрорастающих семян проводили через 48 ч на свету.
Анализ экспрессии QUL1 и QUL2
Экспрессия QUL1 и QUL2 была измерена в мутантах qul1-1 , qul2-2 и qul1-1 qul2-1 , а также в мутантах Col- 0 у 3-недельных растений, выращенных при 22 ° C в течение 16 часов света. РНК выделяли из 20-100 мг розетки растений с помощью набора Invitrap Spin Plant RNA Mini (Strateco) в соответствии с инструкциями производителя.Для синтеза кДНК 1 мкг РНК был преобразован в кДНК с использованием набора для синтеза кДНК iScript (Bio-Rad) в соответствии с инструкциями производителя. Затем кДНК разбавляли до конечного объема 150 мкл для использования в RT-qPCR. Отсутствие геномной ДНК подтверждали сравнением образцов кДНК с образцами РНК, которые не подвергались обратной транскрипции (минус контроль RT). RT-qPCR выполняли на CFX-Connect (Bio-Rad) в 96-луночных планшетах (HSP-9645; Bio-Rad), используя на лунку общий объем 10 мкл, состоящий из 2х.5 мкл кДНК, 0,5 мкл 10 мкМ смеси праймеров, 5 мкл iQ SYBR Green (Bio-Rad) и 2 мкл воды. Для каждой реакции использовалась следующая программа: 95 ° C в течение 3 минут, затем 40 циклов при 95 ° C в течение 15 секунд и 60 ° C в течение 30 секунд с последующим анализом кривой плавления от 65 до 95 ° C. Каждую из четырех биологических повторностей измеряли в двух экземплярах. Экспрессию QUL1 и QUL2 измеряли с использованием праймеров, описанных Fuentes et al. (2010). Для эталонного гена MONENSIN SENSITIVITY1 ( MON1 ; AT2G28390 ) набор праймеров, описанный Chechowski et al.(2005). Данные экспрессии нормализовали с помощью метода 2 -ΔΔ C T (Schmittgen and Livak, 2008).
Статистический анализ
GWA-картирование было выполнено на основе плодородия и архитектурных особенностей, а также времени цветения. GENSTAT (VSN International, 2013) был использован для расчета простых остатков, которые характеризуют воздействие тепла, с поправкой на разницу между образцами в размере силикатов в контрольных условиях.Простые остатки были рассчитаны для средней длины кремнезема в различных участках соцветия, представляющих разные стадии развития. Для расчета простых остатков среднее значение данных, собранных в условиях контроля и теплового стресса, сравнивали с помощью линейной регрессии (наименьшая сумма квадратов), включая все образцы. Картирование GWA было выполнено с помощью программного пакета EmmaX (Kang et al., 2010). Использовалась смешанная модель с поправкой на структуру популяции на основе матрицы родства всех SNP.SNP с частотой минорного аллеля ниже 0,05 были исключены из анализа. Для каждого SNP было рассчитано, какая часть общей фенотипической и генотипической дисперсии может быть объяснена аллелями этого SNP. Коэффициент ранговой корреляции Спирмена (ρ) (двусторонний, α = 0,05) использовался для определения корреляций между рядами данных с помощью SPSS (версия 19.0; IBM Corp.). Односторонний двухвыборочный тест t , предполагающий неравные отклонения, использовался для сравнения среднего числа цветков, образовавшихся на основном соцветии у контрольных и термически обработанных растений.ANOVA и анализ ковариации были использованы для сравнения фенотипических средних для каждого из гаплотипов FLC (P. Li et al., 2014) (дополнительный набор данных 4). Первые 10 основных компонентов матрицы родства использовались в качестве ковариат. Последующие попарные сравнения были выполнены с использованием поправки Бонферрони.
Все последовательности из повторно секвенированных образцов Arabidopsis были получены из 1001genomes.org для 525 образцов, и были загружены файлы 2012 нуклеотидных вариаций по сравнению с Col-0 (TAIR10).Пользовательские сценарии Perl были разработаны для определения позиций с частотой аллелей> 2% (SNP должны быть общими для более чем 11 образцов). Другой сценарий Perl анализирует эти позиции для каждого доступа и выводит либо 1, либо 0 для соответствия или несоответствия Col-0. Полученные данные сохраняются в виде фреймов данных (.csv) на диске. Чтобы вычислить LD, необходимые данные были извлечены из файлов .scv с помощью программы gnu «cut», чтобы вырезать интересующую область. Срезанный кадр данных считывается в R (R Development Core Team, 2012), и по столбцам LD ( r 2 или коэффициент корреляции) может быть определен путем вызова функции R «cor ()» с последующим квадратичным операция.Результаты LD доступны пользователю через веб-интерфейс (только для внутреннего доступа). Пользователь может рассчитать LD в любой области генома.
Номера доступа
Данные о последовательностях из этой статьи можно найти в базах данных Arabidopsis Genome Initiative или GenBank / EMBL под следующими номерами доступа: AT1G13860, AT1G56510, AT1G56580, AT1G56590, AT1G78240, AT2G03G0390, AT2G03490, AT2G034905 AT3G45970, AT3G48190, AT3G48210, AT4G00650, AT4G19710, AT5G10140, AT5G10170, AT5G10200, AT5G10200, AT5G22430 и AT5G22640.
Дополнительные данные
Дополнительный рисунок 1. LD между значимыми SNP, обнаруженными с помощью сопоставления GWA, и окружающими SNP, полученными из данных повторной последовательности.
Дополнительная фигура 2. Экспрессия QUL1 и QUL2 в одинарных и двойных мутантах и Col-0 дикого типа.
Дополнительная таблица 1. Праймеры левой и правой границы, используемые для определения гомозиготности вставок Т-ДНК.
Дополнительный набор данных 1. Даты начала цветения первого экземпляра образца и даты, когда образцы подвергались термообработке.
Дополнительный набор данных 2. SNP, идентифицированных картированием GWA для времени цветения и средней длины кремния в областях развития B, C и D (как определено на рисунке 2) в контрольных условиях и условиях термообработки и их соответствующих остатков. для этих регионов.
Набор дополнительных данных 3. Silique — длина вдоль соцветия подвергшихся тепловому стрессу и контрольных растений нескольких линий вставки Т-ДНК и двух линий с функциональными (FRI-sf2) и нефункциональными (FRI-flc3) аллелями FLC.
Дополнительный набор данных 4. Средняя длина стебля в регионах B, C и D и время цветения для образцов шести наиболее распространенных аллелей FLC (P. Li et al., 2014).
Сетевое функциональное прогнозирование увеличивает генетическую ассоциацию для прогнозирования генов-кандидатов для гиперчувствительности к гистамину у мышей | G3: Гены | Геномы
Abstract
Генетическое картирование — это основной инструмент генетики модельных организмов; однако многие локусы количественных признаков (QTL) содержат десятки или сотни позиционных генов-кандидатов.Приоритет этих генов для проверки часто составляет ad hoc и смещен предыдущими выводами. Здесь мы представляем метод определения приоритетности позиционных кандидатов на основе вычисленной функции гена. В нашем методе используется машинное обучение с функциональными геномными сетями, связи которых кодируют функциональные ассоциации между генами, для выявления сетевых сигнатур функциональной ассоциации с интересующим признаком. Мы демонстрируем метод путем функционального ранжирования позиционных кандидатов в большом локусе на мышах Chr 6 (45.От 9 МБ до 127,8 МБ), связанных с гиперчувствительностью к гистамину (Histh). Histh характеризуется системным истечением сосудов и отеком в ответ на введение гистамина, что может привести к полиорганной недостаточности и смерти. Хотя на риск Histh сильно влияет генетика, мало что известно о его основных молекулярных или генетических причинах из-за генетической и физиологической сложности признака. Чтобы проанализировать эту сложность, мы ранжировали гены в локусе Histh , предсказывая функциональную ассоциацию с множественными процессами, связанными с Histh.Мы объединили эти прогнозы с новыми данными ассоциации однонуклеотидного полиморфизма (SNP), полученными из обзора 23 инбредных линий мышей и данных конгенного картирования. Лучшие гены включали Cxcl12 , Ret , Cacna1c и Cntn3 , все из которых имели сильные функциональные ассоциации и были проксимальнее SNP, расщепляющихся с Histh. Эти результаты демонстрируют возможности сетевых вычислительных методов для определения весьма правдоподобных количественных генов признаков даже в сложных случаях, включающих большой QTL и крайнюю сложность признака.
Идентификация причинных вариантов в локусах количественных признаков (QTL) является центральной проблемой генетики, но генетическое сцепление часто предотвращает сужение QTL до уровня менее нескольких мегабаз (Mb). Таким образом, QTL может содержать сотни генов-кандидатов. Вместо того, чтобы выявить точный ген (или гены), ответственный за вариацию признаков, картирование QTL дает позиционные гены-кандидаты. Строгое сужение QTL путем точного картирования с конгенными штаммами может занять годы или десятилетия, особенно у таких организмов, как мыши, у которых долгое время генерации.Более того, конгенное картирование с высоким разрешением часто показывает, что общий эффект QTL обусловлен множественными сцепленными генами внутри QTL, а не одним геном (Parker et al. 2013; Yazbek et al. 2011). Таким образом, одних только позиционных данных обычно недостаточно, чтобы выдвинуть кандидатуры генов для последующего биологического наблюдения. Чтобы преодолеть ограничения картографических данных, исследователи ищут в QTL вероятные гены-кандидаты. Однако этот отбор обычно осуществляется по специальным критериям с использованием предварительных знаний или литературного поиска.Эта стратегия сильно ориентирована на предварительные знания и очень подвержена ошибкам из-за отсутствия аннотаций. Необходимы строгие и систематические стратегии для различения позиционных генов-кандидатов для механистического наблюдения.
Мы разработали новый подход к ранжированию кандидатов на должность, основанный на функциональной ассоциации с характеристикой. Чтобы избежать аннотаций или предвзятости в литературе, мы используем функциональные геномные сети (FGN), которые кодируют предсказанные функциональные ассоциации между всеми генами в геноме.FGN, такие как Функциональные сети тканей мыши (FNTM) (Goya et al. 2015) и HumanBase (Greene et al. 2015), представляют собой байесовские интеграционные сети, которые объединяют коэкспрессию генов и данные связывания белок-белок. , аннотации онтологии и другие данные для прогнозирования функциональных ассоциаций между генами. С помощью этих сетей мы можем расширить известные ассоциации генов и признаков, чтобы идентифицировать гены, которые ранее не были связаны с этим признаком.
Недавние исследования функциональных геномных сетей продемонстрировали их способность генерировать новые ассоциации между генами и определенными терминами фенотипа (Guan et al. 2010) или биологические процессы (Ju et al. 2013). Например, Guan et al. (2010) использовал классификатор машины опорных векторов (SVM) для идентификации генной сети, связанной с минерализацией костей. Они предсказали и подтвердили новые ассоциации между генами и фенотипами минерализации костей для генов, лежащих за пределами всех опубликованных QTL фенотипов минерализации костей (Guan et al. 2010). Последующие исследования с использованием аналогичных сетевых методов сделали новые прогнозы генов, связанных с гипертонией и аутизмом (Greene et al. 2015; Кришнан и др. 2016). Мы расширили эти методы, чтобы ранжировать гены в картированном QTL на основе множества предполагаемых функциональных терминов и интегрировать эти ранжирования со значениями генетической ассоциации p из исследований штаммов. Мы создали ранжированный список для всех генов в QTL, который включал как функциональные, так и позиционные баллы каждого гена-кандидата.
Наша стратегия сначала построила списки генов, ассоциированных с признаками, из структурированных биологических онтологий ( e.грамм. , Онтология генов (Ashburner et al. 2000; Gene Ontology Consortium 2018) и Онтология фенотипа млекопитающих (Smith and Eppig 2012)) и общедоступные транскриптомные данные из Gene Expression Omnibus (GEO) (Эдгар и др. 2002; Barrett et al. 2012). Затем мы применили классификаторы машинного обучения к функциональным сетям тканей у мышей (FNTM) (Goya et al. 2015), чтобы определить сетевые сигнатуры списков генов, связанных с признаками. Эта стратегия позволила нам предсказать ассоциации ген-признак, которые не были аннотированы в рамках структурированной онтологии, преодолев проблему отсутствия аннотации.
Мы применили наш подход к большому QTL, связанному с гиперчувствительностью к гистамину (Histh) у мышей. Histh у мышей — это летальная реакция на инъекцию гистамина. У нечувствительных мышей инъекция гистамина вызывает воспалительную реакцию, которая проходит без дальнейшего лечения. У мышей с ответом Histh развивается возбуждение и побледнение ушей, за которыми следует прогрессирующий респираторный дистресс, вазодилатация, анафилактический шок и смерть (Vaz et al. 1977; Wang et al. 2014).Histh можно индуцировать в подгруппе линий мышей путем сенсибилизации полным адъювантом Фрейнда (CFA). Histh также развивается спонтанно у животных SJL / J старше шести месяцев.
Ранее мы сопоставили Histh с локусом на Chr 6 (от 45,9 МБ до 127,8 МБ; локус Histh ), что было подтверждено с использованием конгенной линии (B10.S- Histh SJL ) (Raza et al. в печати). Из-за большого размера этого локуса требуется дополнительная информация для выявления причинных вариантов.Чтобы сузить круг кандидатов, мы интегрировали новые данные генетической ассоциации из интервала-специфичных конгенных рекомбинантных линий (ISCRL) и обзор инбредных штаммов с нашими сетевыми функциональными предсказаниями генов, связанных с Histh. Дополнив позиционные данные функциональными прогнозами, мы резко сократили список генов-кандидатов до управляемого набора высококачественных кандидатов, которые участвуют в процессах, связанных с Histh.
Материалы и методы
В качестве дополнения к вычислительной части раздела о методах этот документ включает исполняемый рабочий процесс (см. Доступность данных).Схема вычислительного рабочего процесса показана на рисунке 1. Рабочий процесс включает в себя все файлы и параметры, необходимые для воссоздания вычислительных частей этого исследования.
Рисунок 1Обзор рабочего процесса. Рабочий процесс разбит на блоки по цвету, каждый с заголовком, выделенным жирным шрифтом. Каждый блок показывает, как данные (синие прямоугольники) обрабатывались (серые прямоугольники) для достижения результатов (зеленые прямоугольники). Стрелки показывают общий поток работы и зависимость (и независимость) отдельных анализов.
Животные
Всего 23 линии мышей (129X1 / SvJ, A / J, AKR / J, B10.S- H 2s / SgMcdJ (B10.S), BALB / cJ, BPL / 1J, BPN / 3J, C3H / HeJ, C57BL / 6J, C57BL / 10J, CBA / J, CZECHII / EiJ, DBA / 1J, DBA / 2J, FVB / NJ, JF1 / MsJ, MOLF / EiJ, MRL / MpJ, NOD / ShiLtJ, NU / J, PWD / PhJ, PWK / PhJ, SJL / J и SWR / J были приобретены в лаборатории Джексона ( Бар-Харбор, Мэн). Все мыши, в том числе линии B10.S- Histh SJL и B10.S- Histh SJL ISRC, были созданы и содержались в определенных свободных от патогенов условиях в виварии данного медицинского корпуса. в Университете Вермонта в соответствии с рекомендациями Национального института здравоохранения.Все исследования на животных были одобрены Комитетом по уходу и использованию животных Университета Вермонта.
Histh Фенотипирование
В день 0 мышам вводили 2 подкожно. сайты с полным адъювантом Фрейнда (CFA) (Sigma-Aldrich, Сент-Луис, Миссури) с добавлением 200 мкг г Mycobacterium tuberculosis h47Ra (Difco Laboratories, Детройт, Мичиган). На 30 день определяли гиперчувствительность к гистамину внутривенно. инъекция гистамина (мг / кг сухого веса свободного основания) в фосфатно-солевом буфере (PBS).Смерти регистрировали через 30 минут после инъекции, и данные представляли как количество погибших животных по отношению к количеству исследованных животных. Достоверность наблюдаемых различий определяли по критерию хи-квадрат со значениями p <0,05 достоверно.
Экстракция ДНК и генотипирование
ДНК выделяли из отрезков хвоста мыши, как описано ранее (Sudweeks et al. 1993). Вкратце, отдельные кусочки хвоста инкубировали с буфером для лизиса L-клеток (мкг / мл протеиназы K, 100 мМ NaCl, 10 мМ Трис-HCl (pH 8.3), 10 мМ EDTA, 100 мМ KCl, 0,50% SDS) в течение ночи при температуре. На следующий день добавляли мкл 6 М NaCl с последующим центрифугированием в течение 10 мин при. Слой супернатанта переносили в свежую пробирку, содержащую 1 л изопропанола. После центрифугирования в течение двух минут супернатант удаляли, а осадок промывали 70% этанолом. После последнего двухминутного центрифугирования супернатант удаляли, а ДНК сушили на воздухе и ресуспендировали в L TE.
Генотипирование:
Генотипирование выполняли с использованием микросателлитных маркеров в стандартной реакции ПЦР или специфичных для последовательности праймеров SNP в реакции фототипирования.Полиморфные микросателлиты были выбраны так, чтобы иметь минимальный полиморфизм 8 пар оснований для оптимальной идентификации с помощью электрофореза в агарозном геле. Вкратце, праймеры синтезировали с помощью IDT-ДНК (Коралвилл, ИА) и разбавляли до концентрации М. ПЦР-амплификацию выполняли с использованием Promega GoTaq. Условия цикла включали двухминутный этап начальной денатурации при 94 °, за которым следовали 35 циклов при 94 ° в течение 30 секунд, 55 ° в течение 30 секунд и 72 ° в течение 30 секунд, а затем заключительный этап удлинения при 72 ° в течение пяти минут.Ампликоны подвергали электрофорезу в 2% агарозном геле и визуализировали бромидом этидия и УФ-светом.
Фототипирование:
Генотипирование выполняли с использованием специфичных для последовательности праймеров, которые различаются только 3 ’нуклеотидом, соответствующим каждому аллелю идентифицированного SNP (Bunce et al. 1995). Каждый набор праймеров был разработан с использованием Primer3 с Tm 58-60 ° C, синтезирован с помощью IDT-DNA (Коралвилл, штат Айяо) и использован в концентрации 100 мк M (последовательности праймеров доступны в дополнительном файле 1). .Реакции ПЦР подвергались многостадийным (высокая, средняя и низкая строгость) циклическим условиям, как описано в дополнительном файле 2, и, если было обнаружено, что это необходимо, условия цикла на каждой стадии корректировались для обеспечения оптимальной температуры отжига. Ампликоны подвергали электрофорезу с загрузочным буфером L Orange G на 1,5% агарозном геле, окрашенном бромидом этидия и визуализированном в УФ-свете. Присутствие SNP-специфического аллеля оценивали путем наблюдения ампликона ожидаемого размера в любой реакции.Условия цикла доступны в дополнительном файле 2.
Генерация конгенных линий Histh и GigaMUGA
B10.S- Histh SJL Линии ISRC были созданы путем идентификации рекомбинантных гаплотипов в интервале Histh среди (B10.S- Histh SJL × B10.S) × B10.S BC1, а затем фиксировали в виде гомозиготных линий (рис. 3). Для выявления потенциальных контаминирующих фоновых локусов, сегрегированных среди штаммов, и для дальнейшего уточнения точек разрыва рекомбинации для каждой линии, линии были дополнительно генотипированы с использованием массивов GigaMUGA (143 259 маркеров) коммерческой службой Neogen / Geneseek (Lincoln, NE).
Целевое генетическое ассоциативное тестирование
Мы извлекли данные о генотипах (как кодирующих, так и некодирующих) 23 линий мышей из баз данных Института Сэнгера (Keane et al. 2011) (https: //www.sanger. ac.uk/science/data/mouse-genomes-project) и Лаборатория Джексона (Bogue et al. 2017) (https://phenome.jax.org/). Отсутствие представительства штаммов дикого происхождения , например. , MOLF / EiJ и др. В этих базах данных было компенсировано генотипированием с использованием платформы Illumina Hiseq 2500 https: // www.illumina.com/. Подробные методы см. В дополнительном файле 3. Все эти источники данных были сопоставлены для получения информации о генотипе в общей сложности 15 302 SNP по локусу Histh (45-128 Мбит / с, дополнительный файл 4).
Для расчета ассоциаций между генетическим полиморфизмом и Histh мы использовали эффективную ассоциацию смешанных моделей (EMMA) (Kang et al. 2008). Этот метод рассматривает генетическое родство как случайную величину в линейной смешанной модели для учета структуры популяции, тем самым уменьшая ложные ассоциации между SNP и измеряемым признаком.Для расчета матрицы родства мы составили набор из 470 365 SNP для всех штаммов из базы данных Mouse Genome Informatics (веб-сайт Mouse Genome Informatics Mouse Genome Informatics). Мы удалили SNP в конгеническом интервале из оценки родства и сгенерировали матрицу родства с оставшимися 455 068 SNP. Мы использовали тестовую функцию отношения правдоподобия (emma.ML.LRT) для получения p значений. Значимость определялась поправкой Бонферрони (). Геномные координаты включены для каждого SNP с использованием последней сборки генома мыши GRCm38.p5 / мм10.
Наборы генов, связанных с признаками
Позиционные гены-кандидаты были ранжированы на основе их предсказанной ассоциации с семью функциональными терминами, относящимися к фенотипу Histh: «старение», « Mycobacterium tuberculosis », «сердечная», «связанная с G-белком. рецептор »,« гистамин »,« воспаление »,« гиперчувствительность I типа »и« проницаемость сосудов ». Мы использовали Джина Уивера (Baker et al. 2012) для создания наборов генов, связанных с каждым термином. Для этого мы ввели каждый термин на главную страницу Gene Weaver (https: // geneweaver.org). Мы ограничили поиск генами человека, крысы и мыши и только тщательно подобранными списками. Ортологи мыши для каждого гена были получены с помощью инструмента пакетного запроса в MGI (http://www.informatics.jax.org/batch_data.shtml). Кроме того, мы использовали Gene Expression Omnibus (GEO) и PubMed для получения наборов данных экспрессии для каждого термина фенотипа. Мы удалили всех позиционных кандидатов из списков генов, используемых для обучения, так что для обучения не использовались истинно положительные позиционные кандидаты. Используемые списки генов доступны в виде заархивированных текстовых файлов в дополнительном файле 5.
Сеть FNTM
Мы обучили вспомогательные векторные машины (SVM) классифицировать гены в каждом списке генов с использованием функций, полученных из FNTM (Goya et al. 2015). В этой функциональной геномной сети гены являются узлами, а весовые коэффициенты между ними представляют собой непрерывные значения от 0 до 1, предсказывающие степень, в которой каждая пара генов функционально связана. Более высокие значения указывают на более высокую прогнозируемую функциональную взаимосвязь. Функциональное родство в этой сети было предсказано посредством байесовской интеграции наборов данных из нескольких источников, включая экспрессию генов, данные о межбелковом взаимодействии и аннотацию к терминам GO (Goya et al. 2015). Мы загрузили верхние границы мышиной сети 15 января 2018 г. с http://fntm.princeton.edu.
Кластеризация наборов генов
Guan et al. (2010) заметил, что машины опорных векторов, обученные на 200–300 генах, дают наилучшую точность классификации. В двух из наших списков генов было менее 100 генов. Для всех списков из более чем 400 генов мы уменьшили размер обучающих наборов путем кластеризации каждого набора терминов генов в модули с использованием алгоритма fast greedy (Newman, 2004) в пакете R / igraph (Csardi, 2006).Мы применяли быстрый жадный алгоритм итеративно, пока все модули не содержали менее 400 генов. Использование максимального размера модулей 300 чрезмерно фрагментировало сети, давая много модулей с менее чем 100 генами.
Машинное обучение
Чтобы классифицировать позиционных кандидатов как принадлежащих к функциональному модулю, мы обучили SVM, используя веса соединений в сети FNTM в качестве функций, как описано в Guan et al. (2010). Вкратце, аннотированный набор генов (рис. 2A, синие узлы) используется как набор известных положительных результатов для соответствующего функционального модуля.Ожидается, что другие гены в этом модуле будут прочно функционально связаны с этими известными положительными генами, , то есть , имеют высокую вероятность функционального взаимодействия с известными положительными генами. Таким образом, каждый ген представлен как вектор признаков весов связи с известными положительными результатами, который можно визуализировать как подматрицу матрицы смежности сети (рисунок 2B). Соответственно, строки этой матрицы помечены как заведомо положительные или нет (рис. 2B, синие точки vs. серых точек). Мы использовали пакет e1071 в R (Meyer et al. 2018), чтобы обучить SVM отличать известные положительные гены от набора генов равного размера, выбранных случайным образом вне известного положительного списка с использованием сетевых векторов признаков ( Рисунок 2C). Обученная модель может затем аннотировать новые гены как принадлежащие к функциональному модулю, классифицируя любой ген в геноме (рис. 2C, узлы с зеленой рамкой).
Рисунок 2Сетевое машинное обучение для функциональной аннотации генов.Известные положительные гены, аннотированные функциональным термином (синие узлы), обычно плотно взаимосвязаны в функциональной сети. B Матрица смежности сети — это табличное представление структуры связности сети, в которой каждая строка / столбец соответствует узлу сети, а связанные пары узлов имеют ненулевые значения в соответствующей ячейке матрицы. Обратите внимание, что в целом соединения взвешены, но для отображения мы показываем только существующие / отсутствующие ссылки (белые / черные ячейки).Связи каждого гена в геноме с известными позитивными элементами образуют подматрицу матрицы смежности, называемую матрицей признаков (вертикальные красные линии), строки которой являются векторами признаков для каждого гена. C Используя сетевые векторы признаков для каждого гена, мы обучаем SVM отличать известные положительные (синие точки) от случайных генов в геноме (серые точки), чтобы идентифицировать полную подсеть, соответствующую истинно положительным генам (точки с зеленой рамкой). и пунктирные красные линии на панелях A, B).
Мы обучили 100 SVM каждому модулю, выбирая новый набор случайных генов для каждого прогона.Мы использовали линейное ядро и 10-кратную перекрестную проверку для каждой SVM. Мы обучили каждую SVM по ряду параметров стоимости. Мы начали с последовательности в 10 раз и итеративно сужали диапазон стоимостных параметров, пока не нашли серию из восьми стоимостных параметров, которая максимизировала точность SVM (см. Рабочий процесс).
Мы рассчитали площадь под кривыми ROC (AUC) для всех прогонов следующим образом: для последовательности в диапазоне от минимальной оценки SVM до максимальной оценки SVM мы количественно оценили все истинно положительные (TP), истинно отрицательные (TN) , ложные срабатывания (FP) и ложноотрицательные результаты (FN).Гены TP в этом случае были теми генами из известных положительных результатов, которые были правильно классифицированы как находящиеся в модуле (выше порога оценки SVM). Гены TN в этом случае были те гены вне модуля, которые были правильно классифицированы как находящиеся вне модуля (ниже порога оценки SVM). Мы рассчитали AUC по средней кривой для всех 100 SVM для каждого модуля.
Результаты
Генерация интервальных рекомбинантных конгенных линий (ISRCL) по локусу Histh
В предыдущей работе мы картировали генетический локус, регулирующий чувствительность к гистамину, зависящую от возраста и / или воспаления (CFA), на Chr 6 у мышей SJL / J (Raza et al. в печати). Конгенные мыши B10.S- Histh SJL демонстрировали Histh и несли большую область Mb SJL / J от 45,9 МБ до 127,8 МБ на устойчивом фоне B10.S (Raza et al. In Press) (MGI : 6360897). Этот большой QTL включает 628 генов, кодирующих белок. Чтобы сузить эту область, мы создали пять ISRCL, используя B10.S- Histh SJL x B10.S мышей обратного скрещивания, и оценили их восприимчивость к Histh (рис. 3). В рамках аддитивной модели эти данные предполагают, что Histh состоит из четырех суб-QTL, которые мы обозначили Histh2 (MGI: 6362992), Histh3 (MGI: 6362994), Histh4 (MGI: 6362996), и Histh5 (MGI: 6362997), каждый из которых вносит вклад в общую пенетрантность 17%, 19%, 14% и 10% соответственно.Важно отметить, что для каждого суб-QTL это делает идентификацию гена-кандидата позиционной с использованием интерактивного конгенного картирования с высоким разрешением непрактичной.
Обзор инбредных штаммов Histh
Чтобы выяснить, является ли фенотип Histh уникальным для SJL, мы оценили гистаминовые ответы для 23 инбредных линий мышей (включая SJL / J и B10.S; Таблица 1). Эти штаммы были выбраны с использованием структуры гаплотипов в интервале Histh для идентификации дополнительных линий мышей, которые, вероятно, имеют общий чувствительный аллель Histh (данные не показаны).Мыши 129X1 / SvJ, ALR / LtJ, BPN / 3J, FVB / NJ, NOD / ShiLtJ, NU / J, SJL / BmJ и SWR / J были идентифицированы как имеющие структуру гаплотипа, аналогичную SJL / J в локусе Histh . Мыши ALR / LtJ и SJL / BmJ требовали восстановления эмбрионов и поэтому не были включены в исследование. Фенотипирование Histh идентифицировало мышей FVB / NJ, SWR / J и NU / J как чувствительных к Histh, тогда как 129X1 / SvJ, NOD / ShiLtJ и BPN / 3J были устойчивыми. Взятые вместе с нашими более ранними данными, эти результаты показывают, что восприимчивость к Histh выделяется среди уникального подмножества штаммов, родственных SJL / J (Petkov et al. 2004).
Таблица 1 Обзор фенотипов Histh по 23 инбредным линиям мышейСетевое предсказание ассоциированных с Histh генов
Для прогнозирования функциональных кандидатов среди позиционных кандидатов в локусе Histh мы составили список биологических процессов, связанных с Histh, и обученные классификаторы машинного обучения для определения подсетей функциональных геномных сетей, связанных с каждым из этих процессов. Обзор нашего рабочего процесса представлен на рисунке 1. Сначала мы определили наборы генов, которые были связаны с семью терминами, которые функционально связаны с фенотипом Histh.
Термины и их обоснование следующие:
Гиперчувствительность / анафилаксия I типа : реакция смерти после системной гистаминовой провокации проявляется симптомами гиперчувствительности / анафилаксии I типа, включая респираторный дистресс, вазодилатацию и анафилактический шок (Vaz ). и др. 1977).
Сердечный : есть данные, свидетельствующие о том, что анафилактический шок у мышей связан со снижением сердечного выброса, а не исключительно функцией системной вазодилатации (Wang et al. 2014).
Гистамин : Histh вызывается системным гистамином (Raza et al. In Press).
Рецептор, связанный с G-белком : Гистаминовый рецептор H 1 ( Hrh2 ) передача сигналов требуется для фенотипа Histh, и все рецепторы гистамина принадлежат к семейству рецепторов, сопряженных с G-белком (Hill et al. 1997).
Старение : Спонтанная гистология развивается после шести месяцев у чувствительных линий мышей (Raza et al. в печати).
Воспаление : Лечение провоспалительным CFA вызывает Histh у чувствительных линий мышей.
Tuberculosis : Histh индуцируется у некоторых линий мышей CFA, который содержит инактивированные Mycobacterium tuberculosis (Raza et al. In Press).
Сосудистая проницаемость : реакция Histh включает утечку сосудов в коже и скелетных мышцах согласно оценке Miles’assay (Raza et al. в печати).
Мы использовали Gene Weaver, Gene Expression Omnibus (GEO) и PubMed для получения наборов генов, связанных с каждым из этих терминов (см. Материалы и методы). Наборы генов имели размер от 651 до 1466 генов. Поскольку Guan et al. (2010) обнаружил, что SVM, обученные на наборах генов с примерно 300 генами, лучше всего подходят для сетевого функционального прогнозирования, мы сгруппировали большие наборы генов в модули примерно по 300 генов и проанализировали каждый модуль отдельно (см. Материалы и методы).В дополнительной таблице 4 показано количество генов в каждом модуле, а также пять основных условий обогащения для каждого с использованием пакета R gProfileR (Reimand et al. 2018). Множественные члены этих наборов генов кодируются в локусе Histh . Например, Hrh2 был членом набора генов Anaphylaxis. Чтобы уменьшить систематическую ошибку в классификации, мы удалили все такие гены из каждого набора генов перед обучением SVM. Затем мы обучили ансамбль из 100 SVM каждому набору генов модуля.Мы рассчитали кривые ROC для каждой модели, чтобы количественно оценить способность каждого набора SVM отличать гены внутри набора генов модуля от всех генов вне набора генов модуля. AUC варьировались от 0,9 до 0,975, что указывает на то, что SVM смогли надежно классифицировать гены в каждом списке. Другими словами, каждый набор генов, используемый для определения предполагаемого процесса, связанного с Histh, образует отдельную подсеть полнофункциональной геномной сети. Затем мы применили обученные модели SVM к позиционным генам-кандидатам в локусе Histh .Классифицируя каждого позиционного кандидата, мы можем идентифицировать гены, которые, вероятно, будут функционально связаны с каждым набором генов модуля. Например, для набора генов модуля анафилаксии рецептор гистамина Hrh2 получил положительную оценку, указывающую на то, что SVM предсказывали его принадлежность к набору генов анафилаксии, несмотря на его отсутствие в обучающей выборке. Этот пример представляет собой положительный контроль и показывает, что SVM идентифицируют биологически релевантные паттерны в функциональной геномной сети.Помимо оценки SVM, мы рассчитали количество ложных срабатываний () для каждого гена (см. Материалы и методы). Низкие значения s указывают на высокую степень уверенности в классификации. Детали этого анализа описаны в исполняемом рабочем процессе в качестве дополнения к этому документу (см. Доступность данных).
Интеграция функционального обогащения с генетической ассоциацией
Гены, которые, как предполагается, имеют высокую функциональную связь с признаком, могут не иметь функционально изменчивых аллелей в исследуемой популяции, и поэтому маловероятно, что они будут влиять на наблюдаемые различия штаммов в Histh.Чтобы идентифицировать гены, которые, вероятно, имеют функционально значимые полиморфизмы, мы интегрировали функциональные баллы со значениями SNP-ассоциации p , чтобы сосредоточиться только на тех кандидатах, которые удовлетворяли обоим критериям. Построив максимальный функциональный балл для гена, против (нормализованный до максимальных значений; см. Материалы и методы), мы можем идентифицировать гены, которые, как было предсказано, имеют как высокую функциональную связь с фенотипом Histh, так и, вероятно, имеют функциональные полиморфизмы. которые сегрегированы с восприимчивостью Histh (рис. 5).Синяя линия на рисунке 5 проходит вдоль Парето-фронта набора генов в этом пространстве. Для любого гена этой линии найти ген с более сильной функциональной ассоциацией означает найти ген с более низким значением SNP p и , наоборот, . Гены около фронта Парето имеют либо сегрегационный полиморфизм, либо, как предполагается, функционально связаны с Histh, либо и то, и другое. Все такие гены потенциально являются хорошими кандидатами для экспериментального наблюдения.
Чтобы ранжировать кандидатов с единственной оценкой, мы определили окончательную оценку гена () для каждого гена, которая является суммой (нормализованной) и (рисунок 6).Этот балл отдает предпочтение кандидатам в верхнем правом квадранте с одновременно высокими позиционными и функциональными баллами. Гены в верхнем правом квадранте — Cxcl12 , Ret и Cacna1c — имели почти максимальные баллы по обеим осям и поэтому были признаны лучшими кандидатами для последующего наблюдения. Полную таблицу оценок генов по модулям можно увидеть в дополнительной таблице 2.
В дополнение к идентификации гена с самым высоким рейтингом по полному локусу Histh , мы идентифицировали ген с самым высоким рейтингом для каждого суб-QTL, идентифицированного с помощью конгенных отображение.На рис. 6А показаны функциональные ассоциации по всем модулям 20 лучших генов, отсортированные по окончательной оценке гена (). Полную матрицу оценок для всех ранжированных генов можно найти в дополнительной таблице 2.
Обсуждение
В этом анализе мы идентифицировали небольшой набор позиционных генов-кандидатов в большом локусе путем объединения ассоциаций SNP с предсказанными функциональными ассоциациями с Histh. В результате тестов ассоциации SNP были получены группы SNP со многими значимыми и неразличимыми значениями p .На рис. 5 в правом нижнем углу показана группа генов, которые имеют максимально значимые значения p . Эти гены неотличимы только по величине p , но функциональная оценка в этом конвейере позволяет дифференцировать эти гены на основе родства с интересующими процессами. Более того, множественные гены, получившие высокие оценки по всему конвейеру, не получили максимально значимых значений p из тестов ассоциации SNP. Мы отнесли SNP к генам на основе близости, но это отнесение может вызвать ложные ассоциации между генами и фенотипами.Например, SNP может пометить регуляторный вариант, который влияет на несколько близлежащих генов, только некоторые из которых связаны с этим признаком. Более того, SNP не может полностью отделиться от причинного варианта. Сходным образом сложность генетической архитектуры, включая эпистаз, может приводить к снижению значимости одномерных ассоциаций SNP для истинно причинных SNP. Таким образом, гены с менее оптимальными значениями p можно не принимать во внимание, если значение p было единственным критерием ранжирования.Добавляя функциональный критерий к процессу ранжирования, мы можем защитить от сложностей картирования и ошибок назначения генов и определить, какие из позиционных кандидатов также могут быть функционально связаны с Histh. Окончательный список генов был весьма правдоподобным, и его можно относительно легко проверить с помощью современных методов генетического редактирования.
Высококачественные кандидаты на Histh
Особого внимания заслуживают три гена в итоговом ранжированном списке: Cxcl12 , Ret и Cacna1c .Эти гены не имели наиболее значимых значений p по всему локусу, но, как было предсказано, они были в высокой степени функционально связаны с процессами, связанными с Histh (Рисунок 5). Ген, занимающий первое место, Cxcl12 (также известный как фактор 1, полученный из стромальных клеток (Sdf1)), является хемотаксическим для тучных клеток через хемокиновый рецептор Cxcr4 (Ghannadan et al. 2002). Тучные клетки являются основными движущими силами патологических событий при анафилаксии (Либерман и Гарви, 2016), демонстрируя, что окончательные прогнозы очень важны для Histh.Кроме того, CXCL12 / SDF1, секретируемый опухолевыми клетками, связан с повышенной проницаемостью эндотелия как локально, так и системно. Второй по рангу ген Ret кодирует протоонкоген ret, трансмембранный рецептор клеточной тирозинкиназы, активация которого стимулирует множественные нисходящие пути, участвующие в дифференцировке, росте, миграции и выживании клеток, воспалении (Rusmini et al. ). 2013) и развития сердечно-сосудистой системы (Hiltunen et al. 2000). Вероятно, аллели этого гена могут модифицировать множественные процессы, лежащие в основе Histh, включая как анатомический фон, чувствительный к Histh, так и острую реакцию на гистамин. Ret был значительно связан с множеством функциональных наборов генов (рис. 6А). На третьем месте находится ген Cacna1c , кальциевый канал, зависимый от напряжения, L-тип, альфа-субъединица 1C, который экспрессируется в сердце, кровеносных сосудах и центральной нервной системе (веб-сайт информатики генома мышей).Мутации в Cacna1c связаны с электрофизиологическими изменениями в сердце (Napolitano et al. 2015; Hedley et al. 2009), предполагая возможную роль Cacna1c в нарушении сердечной функции при Histh. Интересно, что SNP в человеческом CACNA1C недавно были связаны с хронической спонтанной крапивницей (, т.е. , спонтанные эпизоды крапивницы и / или ангионевротического отека) и ответом на антигистаминные препараты (Yan et al. 2018).Эти результаты предполагают прямую связь между Cacna1c и гистаминовым ответом.
Все вышеперечисленные гены лежат в локусе Histh5 , который составляет лишь часть общей изменчивости фенотипа Histh. В локусе Histh4 первым был ген-кандидат Cntn3 , который кодирует контактин 3, также называемый белком суперсемейства 1 иммуноглобулинов головного мозга (BIG-1) или нейрональным гликопротеином, ассоциированным с плазмоцитомой (PANG). сообщалось в 1994 г. у крыс (Connelly et al., 1994; Yoshihara et al., 1994). Генетические варианты человеческого CNTN3 связаны с расстройством аутистического спектра (Morrow et al., 2008; Hussman et al., 2011) и сосудистыми аномалиями, лежащими в основе аневризм брюшной аорты (Elmore et al. 2009). Последнее предполагает потенциальную связь с нарушением сердечной функции во время провокации гистамином (Elmore et al. 2009). Интересно, что CNTN3 находится рядом с SNP, разделяющим синдром системной капиллярной утечки (SCLS) из человеческого GWAS.SCLS — чрезвычайно редкое заболевание, характеризующееся преходящими, но потенциально летальными эпизодами диффузной утечки из сосудов белков и жидкостей в периферические ткани, что приводит к массивному отеку всего тела и гипотензивному шоку. Патологические механизмы и генетическая основа SCLS остаются неуловимыми (Xie et al. 2013), но SCLS разделяет многие фенотипические свойства с Histh у мышей. В частности, приступы SCLS диагностируются на основании клинической триады гипотензии, повышенного гематокрита и гипоальбуминемии, все из которых естественным образом возникают у гист-чувствительных мышей линии SJL (Raza et al. в печати). Потенциальная ассоциация между CNTN3 и SCLS, следовательно, также подтверждает его возможную функциональную роль в Histh. Действительно, CNTN3 был не только позиционным кандидатом в SCLS GWAS, но содержался в функциональных терминах, которые были обогащены среди генов-кандидатов наивысшего положения (Xie et al. 2013), что указывает на то, что CNTN3 функционирует совместно с другими генетическими факторы риска SCLS.
В локусе Histh2 наибольшими хитами оказались Creb5 и Tril . Creb5 кодирует белок 5, связывающий цАМФ-чувствительный элемент. Creb5 имеет высокую экспрессию в сердце (Fagerberg et al. 2014) и участвует в сердечной функции и патологии (Schisler et al. 2015). Кроме того, CREB5 является компонентом возрастной воспалительной сети, лежащей в основе нерегулируемой экспрессии цитокинов, то есть воспаления. Tril является взаимодействующим с Tlr4 повторами, богатыми лейцином, и является функциональным компонентом комплекса Tlr4, участвующего в передаче сигналов LPS, и высоко экспрессируется в почках (Carpenter et al. 2009), что указывает на потенциальную роль Tril в регуляции артериального давления. Мыши Tril (- / -) также производят более низкие уровни множественных провоспалительных цитокинов и хемокинов в головном мозге после заражения E. coli и LPS (Wochal et al. 2014) и необходимы для передачи сигналов Tlr3, что предполагает потенциальную возможность роль в иммунной модуляции. В локусе Histh3 значимых совпадений не обнаружено.
Ни один из перечисленных здесь приоритетных генов не был протестирован специально на связь с Histh, и потребуется дальнейшая экспериментальная проверка.Однако каждый из вышеперечисленных генов имеет убедительные функциональные ассоциации, которые могут использоваться в последующих исследованиях.
Вычисление и прогноз гена количественного признака
Окончательная функциональная проверка гена количественного признака (QTG) традиционно требовала либо конгенного картирования для разрешения чрезвычайно узкого QTL, либо ad hoc номинации гена-кандидата для прямого эксперимента. Появление современных генетических технологий, таких как CRISPR / Cas9 (Hsu et al. 2014), позволяют относительно быстрые и недорогие аллельные манипуляции, поэтому бремя предсказания QTG смещается в сторону режима, в котором небольшая горстка сильных кандидатов может отслеживаться индивидуально. Важно отметить, что многие QTL, включая Histh , содержат несколько причинных вариантов, поэтому одно только точное отображение не может обеспечить окончательную проверку. Следовательно, вычислительные инструменты, которые могут идентифицировать небольшое количество разумных кандидатов, могут оказать существенную помощь в биологическом наблюдении. Мы представили интегративную стратегию ранжирования генов в QTL путем объединения предсказанных функциональных ассоциаций с признаком с ассоциациями SNP.Наш метод создает полный ранжированный список генов в локусе, что дает исследователям возможность проверить несколько целей. С этой целью QTL Histh представляет собой крайний вариант использования для прогнозирования QTG — большой полигенный QTL, связанный с физиологически сложным признаком.
Одним из основных ограничений нашего подхода является решение о том, какие функциональные термины включать для сетевого прогнозирования. Чем лучше этот набор приспособлен к интересующей характеристике, тем больше у нас будет уверенности в окончательных прогнозах.В принципе, включение ложного функционального термина может исказить ранжирование в сторону генов, которые функционально связаны с ложным термином, но не имеют отношения к интересующему признаку. Одним из возможных способов решения этой проблемы является использование функциональных данных, таких как транскриптомика, непосредственно от картографической популяции. Однако в некоторых случаях, включая Histh, соответствующая ткань, в которой можно измерить экспрессию гена, может быть неочевидной. В качестве альтернативы можно было бы рассмотреть отдельные рейтинги для каждого функционального термина.В любом случае исследователь должен будет проявить определенную долю суждения в процессе определения приоритетов. Однако, перенеся суждения из большого списка позиционных генов-кандидатов в меньший и более управляемый список биологических процессов, связанных с признаками, мы показали, что можем прийти к сильному набору последующих кандидатов, которые уклонились бы от наивных p значений фильтров и относительно объективны по результатам, опубликованным в литературе.
Еще одним ограничением этого подхода является наш метод присвоения генам значений p .В настоящее время мы используем значение p из единственного лучшего SNP в пределах 1 Мб тела гена, и эти SNP могут или не могут маркировать ген, которому они назначены. Существует несколько методов для присвоения значений p на основе генов, основанных на нескольких значениях SNP p , таких как Универсальный геномный тест для общегеномных ассоциативных исследований (VEGAS) (Liu et al. 2010), Combined Тест ассоциации генов (COMBAT) (Wang et al. 2017) и eigenMT (Davis et al. 2016). Однако ни один из этих методов напрямую не применим к текущему эксперименту, и получение статистики теста на основе генов, которая применима здесь, выходит за рамки данного исследования. Мы будем исследовать эту возможность для будущих реализаций этого метода.
Окончательный результат нашего метода, ранжированный список позиционных генов-кандидатов, легко интерпретируется и предоставляет исследователям четкий набор гипотез для проверки в лаборатории. Хотя этот подход не может окончательно идентифицировать причинный ген или гены в локусе, он предоставляет значительно сокращенный набор вероятных кандидатов для тестирования.
Благодарности
Мы благодарим Лауру Корт за руководство студентами во время генотипирования конгенных мышей. ALT и JMM поддерживаются грантом (R21 LM012615) Национальной медицинской библиотеки Национального института здоровья США (NIH). AR, DNK, EPB и CT были поддержаны грантами NIH и Национального общества рассеянного склероза (NMSS). DNK был поддержан грантами Национального института неврологических заболеваний и инсульта (R01 NS097596), Национального института аллергии и инфекционных заболеваний (R21 AI145306) и NMSS (RR-1602-07780).
- Получено 26 июля 2019 г.
- Принято 20 октября 2019 г.
- Copyright © 2019 Tyler et al.
Это статья в открытом доступе, распространяемая в соответствии с условиями Международной лицензии Creative Commons Attribution 4.0 (http://creativecommons.org/licenses/by/4.0/), которая разрешает неограниченное использование, распространение и воспроизведение в на любом носителе при условии правильного цитирования оригинальной работы.
Цитированная литература
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- 900
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- Meyer,
- Meyer, E. Dimitriadou, K. Hornik, A. Weingessel, and F. Leisch, 2018 e1071: Различные функции Департамента статистики, Группа теории вероятностей (ранее: E1071), TU Wien. Пакет R версии 1.7–0.
- ↵
- ↵
- ↵
- ↵
Raza, A., E.C. Chan, W.-S. Чен, З. Се, Л. М. Скотт, и др. , 2019 Натуральная модель мыши выявляет генетические детерминанты синдрома системной капиллярной утечки (болезнь Кларксона). Биология коммуникации.
- ↵
Рейманд, Дж., Р. Колде и Т. Арак, 2018 gProfileR: Интерфейс к набору инструментов ‘g: Profiler’ . Пакет R версии 0.6.7.
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
Fergus Ewing для компании LOW IN и MSPATING
Правительство Шотландии выделило 1,75 миллиона фунтов стерлингов на три инновационных проекта в области отопления.
В схемах на Шетланде, Клайдбанке и Глазго будет использоваться технология теплового насоса с водяным источником для извлечения тепла из воды — даже в самые холодные дни — для эффективного снабжения низкоуглеродным теплом.
Это:
- Кредит в размере 1,6 миллиона фунтов стерлингов для крупномасштабной схемы теплового насоса с источником морской воды в Леруике, чтобы позволить еще 225 домохозяйствам присоединиться к существующей тепловой сети
- Финансирование в размере 75 000 фунтов стерлингов для развития Queens Quay на территории бывшей верфи Джона Брауна для разработки инвестиционного проспекта для сети централизованного теплоснабжения с использованием водяного теплового насоса в бассейне реки Клайд.
- Финансирование в размере 75 000 фунтов стерлингов для Западного кампуса Университета Глазго для разработки предложения инвестиционного уровня по установке водяного теплового насоса на реке Кельвин, чтобы существующая сеть централизованного теплоснабжения могла обслуживать новые здания, запланированные на месте бывшей больницы Western Infirmary .
Министр энергетики Фергус Юинг сделал заявление сегодня на ежегодной конференции Scottish Renewables в Эдинбурге.
Г-н Юинг сказал:
«Поддержка развития централизованного теплоснабжения и более широких низкоуглеродных технологий поможет максимально использовать экономические возможности низкоуглеродного сектора Шотландии.
«По оценкам, на тепло приходится более половины общего энергопотребления Шотландии и почти половина выбросов парниковых газов в Шотландии, поэтому необходимость принятия мер совершенно очевидна.
«Непрерывный рост числа домов и предприятий, получающих выгоду от подключения к низкоуглеродному доступному теплу, обеспечиваемому сетями централизованного теплоснабжения, помогает правительству Шотландии реализовать наши амбиции по увеличению количества подключений к сетям централизованного теплоснабжения к 2020 году».
Дэйв Пирсон, директор Star Renewable Energy сказал:
«Star приветствует сегодняшнее объявление о том, что правительство Шотландии поддерживает два проекта по разработке предложений инвестиционного уровня по развертыванию тепловых насосов с водяным источником тепла.Наша компания уже несколько лет находится в авангарде использования больших тепловых насосов. В 2009 году мы приняли вызов, поставленный норвежским городом Драммен, по развертыванию большого водяного теплового насоса, собирающего тепло из фьорда для использования по всему городу в их системе централизованного теплоснабжения.
«Компания Drammen сократила свой углеродный след и выбросы из дымовой трубы более чем на 80 процентов, перейдя со сжигания газа на наш тепловой насос с температурой 90 ° C, достигнув целей COP21 2050, заявленных в Париже. Обязательство правительства Шотландии станет катализатором аналогичного шага в Шотландии, поскольку мы стремимся к значительной декарбонизации отопления.”
Тысячи домов будут отапливаться с помощью первой в своем роде системы «poo-power»
Вскоре тысячи домов можно будет отапливать за счет бытовых отходов с помощью первой в своем роде системы «poo-power».
Thames Water планирует использовать избыточное тепло, рекуперированное в процессе очистки сточных вод, для обогрева более 2000 новых домов в Кингстоне, на юго-западе Лондона, в рамках плана регенерации.
Вместо того, чтобы возвращать всю прошедшую окончательную обработку теплую воду непосредственно в реку, около одной трети будет улавливаться и отправляться в новый энергетический центр на территории компании Hogsmill, где она поможет нагреть воду, подключенную к жилому комплексу Kingston Cambridge Road. .
В случае успеха до семи гигаватт-часов низкоуглеродного тепла в год можно было бы поставлять через герметичную сеть труб в систему централизованного теплоснабжения в поместье, когда она в конечном итоге будет восстановлена.
Thames Water планирует затем расширить сеть, включив в нее общественные и коммерческие здания в центре города Кингстон.
Подробнее
Эта схема является первой в своем роде в Англии, но есть надежда, что она послужит моделью для аналогичных проектов в других местах Великобритании, сократив миллионы тонн выбросов углерода.
Thames Water заявила, что проект в Hogsmill позволит сэкономить 105 000 тонн выбросов в эквиваленте углекислого газа за 30 лет — столько же, сколько около 157 000 обратных рейсов из Лондона в Нью-Йорк или более 15 000 поездок на автомобиле по всему миру.
Подпишитесь на новости и советы по спасению планеты
Планируется, что будущие фазы сети централизованного теплоснабжения позволят сократить дополнительные выбросы и помогут Совету Кингстона достичь своей цели по снижению выбросов углерода к 2038 году.
(Вода Темзы)
Сара Бентли, исполнительный директор Thames Water, сказал: «Возобновляемое тепло из нашей канализационной сети — фантастический ресурс, поэтому жизненно важно, чтобы мы были ведущим игроком в энергетическом переходе и раскрыли весь потенциал« энергии пу ».
«Защита и улучшение окружающей среды чрезвычайно важны для нас, и мы взяли на себя обязательство сделать все возможное, чтобы найти новые и инновационные способы достижения наших чистых нулевых амбиций в течение следующих 10 лет.
«Мы уже самостоятельно генерируем значительное количество возобновляемой энергии на нашем огромном участке, удовлетворяя около четверти наших общих потребностей в электроэнергии, и уверены, что инновационные схемы централизованного теплоснабжения, подобные этой, предложат гораздо больше возможностей, чтобы гарантировать, что мы покинем нашу планету. в лучшем месте для будущих поколений.
Глава Совета Кингстона Кэролайн Керр добавила: «Это новаторский подход. Это впервые в Англии и показывает, что мы серьезно относимся к сокращению выбросов углерода в районе. Это реальная возможность проявить смелость и амбициозность для будущих поколений. Приятно работать вместе с Thames Water, превращая отходы в чистую энергию ».
Новости Великобритании в картинках
Показать все 501/50 Новости Великобритании в картинках
Новости Великобритании в картинках
28 февраля 2021 года
Люди гуляют по пляжу в Скарборо, Северный Йоркшир
PA
Новости Великобритании в картинках
27 февраля 2021 года
Гроб капитана Тома Мура несут члены Вооруженные силы в крематории Бедфорда в Бедфорде
EPA
Новости Великобритании в фотографиях
26 февраля 2021 года
Собака стоит на скалах, когда солнце встает позади нее на пляже Нью-Брайтон, Wirral
PA
Новости Великобритании в фотографиях
25 февраля 2021 года
Теневой канцлер Аннелиз Доддс выступает перед бюджетом Института глобального процветания в штаб-квартире Лейбористской партии в Лондоне
PA
Новости Великобритании в картинках
24 февраля 2021 года
Министр здравоохранения Великобритании Мэтт Хэнкок выглядит на улице Даунинг-стрит в Лондоне, Великобритания
REUTERS
Новости Великобритании в картинках
23 февраля 20 21
Два серфера пробираются в море у пляжа Боскомб в Дорсете
PA
Новости Великобритании в фотографиях
22 февраля 2021 года
Крокусы цветут возле Тринити-колледжа в Кембридже
PA
Новости Великобритании в картинках
21 февраля 2021 года
Стрит-арт украшает многие улицы и стены вокруг района Дигбет в Бирмингеме
PA
Новости Великобритании в фотографиях
20 февраля 2021 года
Части Калландера в Стирлингшире затоплены после того, как река Тейт вышла из берегов
PA
Новости Великобритании в картинках
19 февраля 2021 года
Солнце встает над маяком Святой Марии, Нортумберленд
PA
Новости Великобритании в картинках
18 февраля 2021 года
Автомобили едут по трассе A1101 в Уэлни, Норфолк , который снова становится проходимым после двух месяцев затопления паводковыми водами
PA
Новости Великобритании в фотографиях
17 Февраль 2021 г.
Профессор Грин объявляет, что новый 25-километровый суперканал Tideway «направляется на восток», знаменуя собой важный этап в строительстве туннеля, поскольку две гигантские бурильные машины туннеля соединяются, чтобы прорваться в Бермондси и начать прокладку туннеля в направлении Восточного Лондона.После завершения канализационная система предотвратит попадание миллионов тонн неочищенных сточных вод в Темзу
PA
Новости Великобритании в картинках
16 февраля 2021 года
человек держит таблички с надписью «Я ПОТЕРЯНУ СВОЮ РАБОТУ !!» и «БОРИС, КТО ОПЛАТИТ ЭТОТ СЧЕТ? У МЕНЯ НЕТ ДЕНЕГ ДЛЯ ОПЛАТЫ КРЕДИТНОЙ КАРТОЙ »в окне его гостиничного номера в отеле Radisson Blu в Лондоне
Getty
Новости Великобритании в картинках
15 февраля 2021 года
Пассажиры, прибывающие в Терминал 5 аэропорта Хитроу: в сопровождении сотрудников службы безопасности до автобусов.С сегодняшнего дня люди, прибывающие из 33 стран «красного списка», в том числе из ЮАР и Объединенных Арабских Эмиратов, должны изолироваться в номерах отелей на 10 дней за свой счет. Политика была объявлена в конце прошлого месяца в ответ на появление новых вариантов нового коронавируса, которые более устойчивы к существующим вакцинам.
Getty
Новости Великобритании в картинках
14 февраля 2021 года
Люди играют в хоккей на замерзших затопленных полях возле Эли в Кембриджшире
PA
Новости Великобритании в картинках
13 февраля 2021 года
в костюме премьер-министра Бориса Джонсона, маска для лица в сопровождении Сары Роуз (слева), доктора медицинских наук центра по производству средств индивидуальной защиты в Нортумбрии в Ситоне-Делавале
PA
Новости Великобритании в фотографиях
12 февраля 2021 года
Фонари висят на улице, чтобы отпраздновать китайский лунный Новый год Год, который отмечает Год Быка в китайском квартале в центре Лондона, во время третьего национального карантина в Англии для сдерживания распространения коронавируса.
PA
Новости Великобритании в фотографиях
11 февраля 2021 года
Вице-президент Европейской комиссии Марош Сефкович на вокзале Сент-Панкрас в Лондоне после прибытия в Великобританию перед переговорами с министром кабинета министров Майклом Гоу по протоколу Северной Ирландии.
PA
Новости Великобритании в картинках
10 февраля 2021 года
Замерзший дорожный знак и живая изгородь покрыты сосульками, поскольку шторм Дарси поражает большую часть страны, в Шенли, Хартфордшир
Reuters
Новости Великобритании в картинках
9 февраля 2021 года
Люди идут по Гарднер-стрит в Глазго, когда снег покрывает город
AFP / Getty
Новости Великобритании в картинках
8 февраля 2021 года
Снег укрывает автомобиль в Харвуде, графство Дарем
PA
Новости Великобритании в картинках
7 февраля 2021 года
Люди идут по снегу в парке Ноле в Севеноуксе
Getty
Новости Великобритании в картинках
6 февраля 2021 года
Мальчик катается на санях в Мулен-Мур возле Питлохри, как шторм Дарси приближается, в Шотландии
Reuters
Новости Великобритании в фотографиях
5 февраля 2021 года
Ассистент исследовательского центра Лайла Хиллсде n с редкой копией фолио Бомонта и Флетчера 1647 года, антологии пьес Джона Флетчера и Уильяма Бомонта, из личной библиотеки короля Карла II, которая недавно была приобретена Специальными коллекциями в Университете Лидса
PA
Новости Великобритании в фотографиях
4 февраля 2021 года
Эл Гудридж из Ньютонмора использует топор, чтобы создать канал во льду, чтобы его жена Алиса могла плавать в Лох-Инше, в национальном парке Кэрнгормс
PA
Новости Великобритании в картинках
3 февраля 2021 г.
Фреска с изображением капитана сэра Тома Мура работы художника Роберта Ньюбиггина украшает стену в Саутпорте.Ветеран Второй мировой войны собрал почти 33 миллиона фунтов стерлингов для благотворительных организаций NHS в преддверии своего 100-летия в прошлом году, прогуливаясь по своему саду в Марстон-Мортейне, Бедфордшир. Он был госпитализирован в больницу Бедфорд в эти выходные после положительного результата теста на Covid-19 и вчера умер в присутствии своей семьи
Getty
Новости Великобритании в фотографиях
2 февраля 2021 года
Полицейские удаляют активиста-эколога, протестующего против высокой скорости HS2. железнодорожная ветка с крыши офиса HS2 в центре Лондона 2 февраля 2021 года.- Разработанный для работы в бывшем промышленном центре Бирмингема, а затем в Манчестере и Лидсе, HS2 должен был следовать от южного лондонского соединения Eurostar с Парижем
AFP через Getty Images
Новости Великобритании в фотографиях
1 февраля 2021 года
Кристин Барри поднимает двухлетнего сына Орана, чтобы поцеловать статую святой Бригитты, когда они посещают святой колодец святой Бригитты в Ко Килдэр, чтобы отметить День святой Бригитты, который многие в Ирландии считают первым днем весны
PA
Новости Великобритании на фотографиях
31 января 2021 года
Люди выгуливают собаку в парке Pugneys Country Park в Уэйкфилде, Западный Йоркшир
PA
Новости Великобритании в фотографиях
30 января 2021 года
Пожарные осматривают обугленные остатки квартала в Napier Barracks после Полиция заявила, что в Фолкстоне, Англия, был преднамеренно начат пожар.Казармы Нэпир, часть заброшенных казарм Сомерсет-хауса сэра Джона Мура, с лета прошлого года использовались для размещения просителей убежища, прибывающих из Франции на Южное побережье. Сообщается, что 120 из 400 человек, размещенных там, заразились коронавирусом, и приходящий врач описал условия жизни как «бесчеловечные»
Getty
Новости Великобритании в фотографиях
29 января 2021 года
Члены мусульман-ахмадийцев Община принимает участие в пятничной молитве в мечети Байтус Субхан в Кройдоне, Англия.В течение почти последнего года пандемия Covid-19 и связанное с ней ограничение общественных собраний вынудили религиозные общины искать новые способы соблюдения молитв, при этом многие события отмечались дома. Мечети, как и другие места религиозного поклонения, остаются открытыми, хотя их количество было ограничено, чтобы поддерживать социальное дистанцирование
Getty
Новости Великобритании в картинках
28 января 2021 года
Художественные работы с изображением животных, созданные Раймондом и Леонардом, показаны на окно дома в Актоне, Лондон, для открытия The Great Big Art Exhibition, крупнейшей в стране выставки, по инициативе Firstsite
PA
Новости Великобритании в фотографиях
27 января 2021 года
Рыбацкие лодки, некоторые из них заброшены, лежат в водах реки Уайр в Флитвуде в Ланкашире
PA
Новости Великобритании в фотографиях
26 января 2021 года
Группа друзей катится на санях в заснеженном парке Lickey Hills Country Park, Бирмингем
PA
Новости Великобритании в картинках
25 января 2021 года
Велосипедист наблюдает за восходом солнца с Примроуз-Хилл
PA
Новости Великобритании на фотографиях
24 Janaury 2021
Люди катаются на санях по снегу на Парламентском холме в Хэмпстед-Хит
Getty
Новости Великобритании в фотографиях
23 января 2021 года
Дениз Мандей (слева) и Рут Ричардс (справа) проверяют, как Спасенные бывшие куры Рут в паводке в ее доме недалеко от Набурн-шлюза в Йорке после сильного дождя во время шторма Кристоф
PA
Новости Великобритании в фотографиях
22 января 2021 года
Очистка, поскольку паводковая вода начинает отступать от город Нортвич, Чешир, после шторма Кристоф
Getty
Новости Великобритании в фотографиях
21 января 2021 года
Члены службы экстренной помощи работают над эвакуацией жителей Дома престарелых после того, как они оказались в результате наводнения, в Нортвиче, северо-западная Англия, поскольку шторм Кристоф приносит проливные дожди и наводнения по всей Англии
AFP via Getty
Новости Великобритании в картинках
20 января 2021
Наводнение под трассой A46 на Six Hills Lane в Лестершире, поскольку шторм Кристоф собирается принести масштабные наводнения, ураганы и снегопады в некоторых частях Великобритании
PA
Новости Великобритании в фотографиях
19 января 2021 года
Серфер оседлал волну в море у пляжа Боскомб в Дорсете
PA
Новости Великобритании в фотографиях
18 января 2021 года
Премьер-министр Великобритании Борис Джонсон стоит у комнаты, где техники производят вакцину AstraZeneca covid-19 в Oxford Biomedica , в Оксфорде
Reuters
Новости Великобритании в картинках
17 января 2021 года
Люди на берегу моря в Брайтоне во время третьей национальной блокировки Англии для сдерживания распространения коронавируса
PA
Новости Великобритании в картинках
16 января 2021
В соборе Солсбери, графство Уилтшир, установлены отсеки для людей, которым вводят вакцину от коронавируса Pfizer. e
PA
Новости Великобритании в картинках
15 января 2021 года
Транспортные средства проезжают по затопленной дороге в Маунтсоррел, Лестершир, с большой полосой Англии, охваченной 30 предупреждениями о наводнении к обеду в пятницу
PA
Новости Великобритании в картинках
14 января 2021 года
работников NHS из больниц по всей столице во время социально дистанцированной акции протеста за пределами Даунинг-стрит в Лондоне, приуроченной к еженедельным аплодисментам для героев
PA
Новости Великобритании в картинках
13 января 2021 года
Уведомление общественного здравоохранения о необходимости «оставаться дома» на автобусной остановке в Лондоне
EPA
Новости Великобритании в картинках
12 января 2021 года
Солнце встает за замком Скарборо в Йоркшире
PA
Новости Великобритании в картинках
11 января 2021 г.
Медицинский работник наполняет шприц вакциной против Covid-19 в центре вакцины NHS, который был открыт в ЦБ.